組蛋白修飾檢測
組蛋白后轉錄修飾(PTMs),也簡稱為組蛋白修飾,包括甲基化、乙酰化、磷酸化等,對于染色體包裝、轉錄調控以及DNA損傷和修復等各種生物活動起著重要作用 [1-4]。因此,準確敏感地檢測組蛋白修飾對于理解生物過程的表觀遺傳調控和組蛋白修飾酶(HME)靶向藥物的開發具有重要意義。
組蛋白修飾檢測在揭示表觀遺傳調控的關鍵機制中起著重要作用。通過了解組蛋白修飾的重要性、原理和檢測方法,我們可以更好地理解組蛋白修飾在基因調控、發育和疾病中的關鍵機制。
1. 組蛋白修飾檢測方法
組蛋白修飾分析是一系列實驗方法,用于分析和確定組蛋白的修飾狀態和位置,主要包括染色質免疫沉淀(CHIP)和質譜(MS)。這些方法利用特定的實驗策略和工具,檢測帶有特定修飾的組蛋白分子,并定量確定它們的存在量和分布。
1.1 染色質免疫沉淀實驗(ChIP)
ChIP是一種常用的組蛋白修飾檢測技術,用于檢測修飾的存在及其與基因表達的關聯。該技術使用特異性抗體結合修飾的組蛋白,將修飾的組蛋白從細胞或組織中富集出來。然后,可以通過PCR、測序或質譜等方法對富集的修飾組蛋白進行進一步分析,確定修飾的存在和定位。
在檢測組蛋白修飾方面,有三種主要的基于ChIP的方法,包括ChIP-chip、ChIP-SAGE和ChIP-Seq。
在ChIP中,有兩種類型的ChIP:天然ChIP和交聯ChIP。天然ChIP使用通過核酸酶消化細胞核制備的天然染色質。交聯ChIP使用甲醛固定的染色質,并通過聲波破碎進行斷裂。在表1中,我們對比了這兩種ChIP方法。
表1: 天然ChIP和交聯ChIP的比較
| 類型 | 類型 | 劣勢 |
|---|---|---|
| 天然ChIP |
|
|
| 交聯ChIP |
|
|
此外,您可以點擊以下鏈接查看ChIP操作具體流程:
1.1.1 ChIP-chip
ChIP-chip(染色質免疫沉淀結合DNA微陣列雜交)是一種將染色質免疫沉淀與DNA微陣列相結合的技術。與常規的ChIP一樣,ChIP-chip用于研究體內蛋白質與DNA的相互作用。實際上,ChIP-chip方法可用于研究許多表觀基因組現象。這里出現的示例展示了ChIP-chip在研究組蛋白修飾中的應用。正如圖1所示:
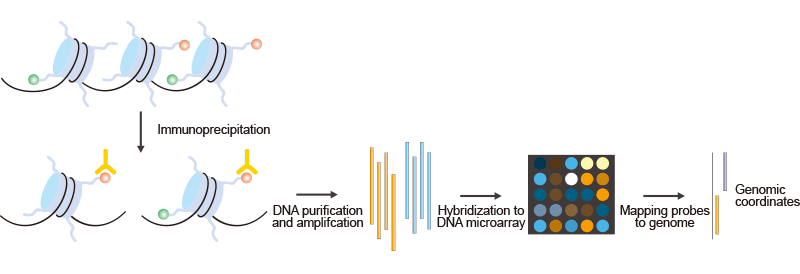
圖1. ChIP-on-chip的簡要步驟
首先,通過使用特定于特定組蛋白修飾的抗體免疫沉淀交聯染色質來純化修飾的染色質。然后,DNA被擴增以獲得足夠的DNA(用一種顏色標記)。用顏色標記的ChIP DNA與從輸入染色質中準備的控制DNA混合,并用不同顏色標記。隨后,微陣列探針可以映射到基因組上,得出基因組坐標。然而,大多數ChIP-on-chip實驗仍然使用多克隆抗體,這些抗體在批次之間的特異性不同。因此,生成和使用單克隆抗體進行ChIP-on-chip實驗是一個重要目標。
1.1.2 ChIP-SAGE
ChIP-SAGE是指染色質免疫沉淀結合串聯分析基因表達。將ChIP實驗與SAGE結合,可以在基因組范圍內描述組蛋白修飾。ChIP-SAGE過程始于ChIP步驟,用于純化與特定組蛋白修飾相關的染色質區域,然后按照以下步驟進行。如圖2所示:
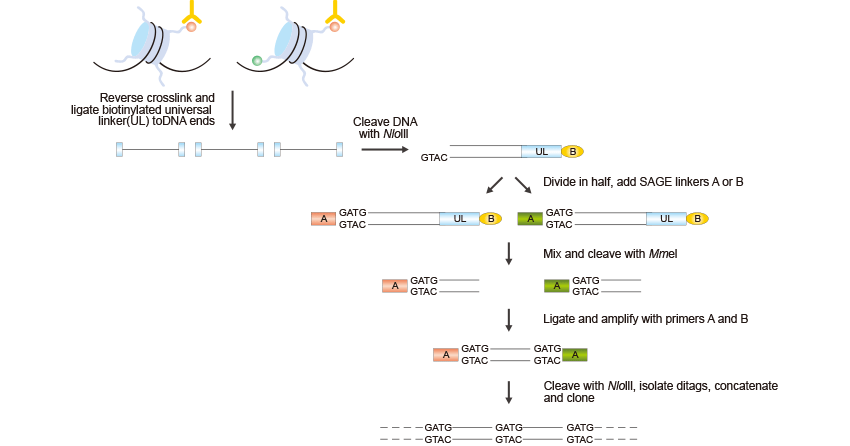
圖2. ChIP-SAGE的簡要步驟
首先,交聯被逆轉,接著將生物素化的通用連接器(UL)連接到DNA末端,并將DNA結合到鏈霉親和素珠上。然后,使用識別CATG的Nla III酶消化DNA,將包含Mme I識別序列的連接器連接到切割的DNA末端。Mme I酶消化產生來自免疫沉淀片段的21-22 bp序列標簽;這些序列標簽被連接在一起,克隆到測序載體中,然后進行測序。每個測序反應中通常可以生成大約20到30個21 bp的短序列標簽。然后,這些序列標簽可以映射到基因組上,以識別修飾區域。
1.1.3 ChIP-Seq
ChIP-seq(染色質免疫沉淀結合高通量測序技術)是近年來在基因組尺度上研究表觀遺傳現象的最令人興奮的技術之一,它依賴于將ChIP實驗與高通量測序技術相結合。ChIP-seq將ChIP與高通量DNA測序相結合,用于鑒定DNA相關蛋白的結合位點。如圖3所示:
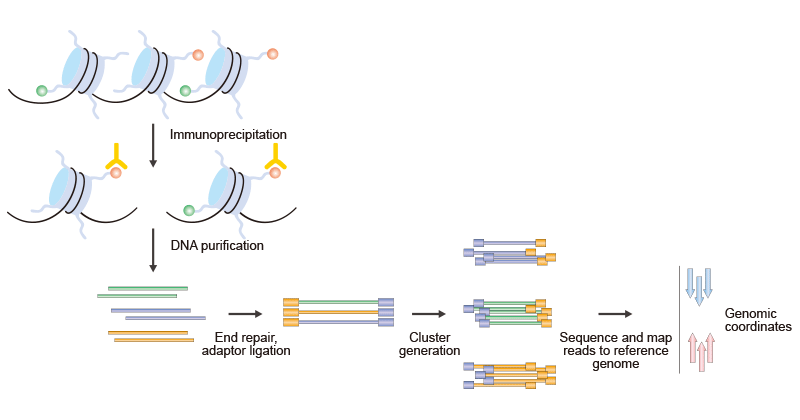
圖3. ChIP-Seq的簡要步驟
這里概述的過程是針對使用Illumina Genome Analyzer和Solexa技術的。它可以分為兩個部分,ChIP和測序。第一步是使用特定于特定組蛋白修飾的抗體對修飾的染色質進行免疫沉淀。然后,修飾的ChIP DNA末端經修復,并與一對適配器連接,隨后進行有限的PCR擴增。DNA分子結合到一個包含共價結合的寡核苷酸的流動細胞表面,這些核苷酸可以識別適配器序列。經過尺寸選擇,所有產生的ChIP-DNA片段都可以使用基因組測序儀同時進行測序。得到的序列讀數被映射到參考基因組上,以獲得與免疫沉淀片段對應的基因組坐標。
1.2 質譜法(MS)
質譜法近年來已成為鑒定和定量組蛋白修飾的首選方法。通過質譜法鑒定和定位特定殘基的組蛋白修飾依賴于肽或蛋白質的實驗測量質量和預期質量之間的"質量差"。理論上,這種方法使得在單次分析中可以對任何PTM或PTM組合進行概要分析,而不受PTM類型或其特定位點的限制,并提供高度準確的定量數據。
質譜法已被證明是評估和比較不同樣本中特定修飾及其組合水平的有價值工具。
在生物樣本中,有三種基于質譜法的方法可用于檢測組蛋白修飾,包括“自上而下”、“中間范圍”和“自下而上”的質譜法 [5-8]。
1.2.1 “自上而下”方法
在“自上而下”方法中,完整的組蛋白被色譜分離,然后被離子化并進行質譜分析 [5,8]。由于完整組蛋白的高電荷狀態,可以從13+到25+不等,所以高分辨率的質譜和質譜/質譜對于這種技術至關重要。這個過程提供了在給定樣本中所有組蛋白同工型以及它們的總體化學計量學的詳細數據。
1.2.2 “中間范圍”方法
在“中間范圍”方法中,通過Glu-C或Asp-N的消化生成長的組蛋白肽,通常分子量超過5 kDa(>20個氨基酸)[5,8]。通過這種方法,可以獲得完整的N-末端尾巴。例如,Asp-N產生H4 1–24肽,而Glu-C產生H3 1–50肽,其中包括了H4和H3中大多數著名PTM位點。盡管這兩種方法適合于檢測長程PTM關聯并估計其豐度,但是這兩種方法面臨的一個主要困難是區分異構肽(具有相同修飾但位于不同位置的肽段),此外,這些方法通常面臨靈敏度降低和計算密集型數據分析的困擾。因此,“自上而下”和“中間范圍”質譜法通常只在少數專業實驗室中進行,并且截至目前尚未有在臨床樣本中應用的記錄。
1.2.3 “自下而上”方法
在“自下而上”質譜法中,組蛋白在通過質譜分析之前,經過Arg‐C蛋白酶介導的酶促消化為相對較短的肽(長度為5-20個氨基酸)。由于核心組蛋白富含堿性氨基酸殘基,胰蛋白酶產生的肽太短,并且在修飾位點附近低效切割,導致長度不一致,不適合準確定量。在胰蛋白酶切割之前,通過氘化乙酸酐或丙酸酐在N-末端和賴氨酸上的游離胺衍生通常用于模擬Arg-C消化,同時使用強大的胰蛋白酶 [9-12]。這種策略也有助于區分等壓肽 [10]。
雖然“自下而上”提供有限的同時發生的PTM數據(最多四個),特別是對于遠處的標記,但它提供了靈活性,并在分析患者來源的樣本中具有應用。
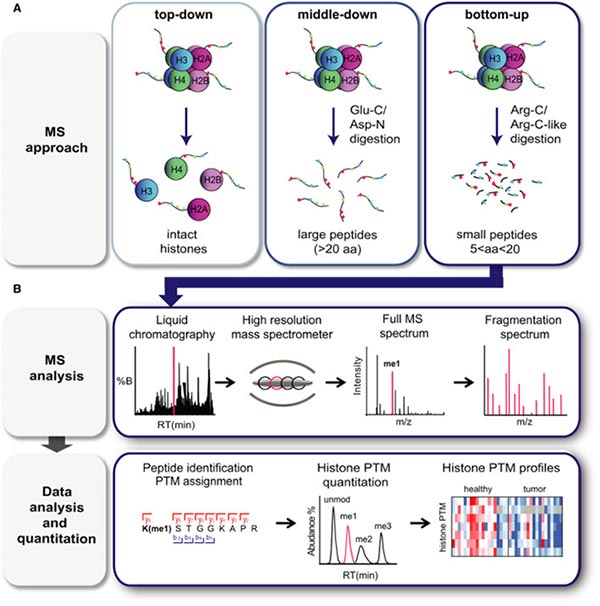
圖4. 基于質譜法的組蛋白修飾分析
圖片引用自: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9291046/
2. 組蛋白修飾檢測的應用
組蛋白修飾檢測在表觀遺傳學研究和個體化醫學中至關重要。它有助于理解基因調控機制,探索細胞分化和組織發育過程,識別疾病生物標志物,并開發靶向治療方法。
2.1 揭示基因調控機制
組蛋白修飾檢測可以幫助我們揭示基因調控機制。通過分析基因啟動子和增強子中不同修飾的分布,我們可以了解組蛋白修飾在基因表達中的調節作用,并進一步了解基因調控的詳細機制。
2.2 研究發育和細胞命運決定
組蛋白修飾檢測在發育過程和細胞命運決定中發揮重要作用。通過分析修飾的動態變化,我們可以研究細胞分化和組織發育的分子機制,以及干細胞命運的調控。
2.3 探索疾病發病機制
組蛋白修飾的檢測幫助我們了解疾病的機制。不同疾病中異常的組蛋白修飾模式可以為疾病的發生和發展提供重要線索,從而為疾病的診斷和治療提供新的靶標和策略。
參考文獻:
[1] Valensisi, C., Liao, J.L., Andrus, C. et al. cChIP-seq: a robust small-scale method for investigation of histone modifications [J]. BMC Genomics 16, 1083 (2015).
[2] Peterson CL, Laniel MA. Histones and histone modifications [J]. Curr Biol. 2004;14(14):R546–51.
[3] Bannister AJ, Kouzarides T. Regulation of chromatin by histone modifications [J]. Cell Res. 2011;21(3):381–95.
[4] Binder H, Steiner L, et al. Transcriptional regulation by histone modifications: towards a theory of chromatin re-organization during stem cell differentiation [J]. Phys Biol. 2013;10(2):026006.
[5] O’Geen H, Echipare L, Farnham PJ. Using ChIP-Seq technology to generate high-resolution profiles of histone modifications [J]. Methods Mol Biol. 2011;791:265–86.
[6] Noberini R, Robusti G, Bonaldi T. Mass spectrometry-based characterization of histones in clinical samples: applications, progress, and challenges [J]. FEBS J. 2022 Mar;289(5):1191-1213.
[7] S. Sidoli, B.A. Garcia. Characterization of individual histone posttranslational modifications and their combinatorial patterns by mass spectrometry-based proteomics strategies [J]. Methods Mol. Biol., 1528 (2017), pp. 121-148.
[8] Z.-F. Yuan, A.M. Arnaudo, B.A. Garcia. Mass spectrometric analysis of histone proteoforms [J]. Annu. Rev. Anal. Chem., 7 (2014), pp. 113-128.
[9] A. Moradian, A. Kalli, M.J. Sweredoski, S. Hess. The top-down, middle-down, and bottom-up mass spectrometry approaches for characterization of histone variants and their post-translational modifications [J]. Proteomics, 14 (2014), pp. 489-497.
[10] Smith CM, Haimberger ZW, et al. (2002) Heritable chromatin structure: mapping "memory" in histones H3 and H4 [J]. Proc Natl Acad Sci USA 99 (Suppl 4), 16454–16461.
[11] Soldi M, Cuomo A & Bonaldi T (2014) Improved bottom‐up strategy to efficiently separate hypermodified histone peptides through ultra‐HPLC separation on a bench top Orbitrap instrument [J]. Proteomics 14, 2212–2225.
[12] Sidoli S, Bhanu NV, Karch KR, Wang X & Garcia BA (2016) Complete workflow for analysis of histone post‐translational modifications using bottom‐up mass spectrometry: from histone extraction to data analysis [J]. J Vis Exp 111, 54112.





