組蛋白變異體
1977年,Franklin和Zweidler首次使用酸-尿素-三聯物X(AUT)聚丙烯酰胺凝膠電泳從哺乳動物組織中分離出組蛋白變異體 [1]。組蛋白具有廣泛的功能,包括基因表達、染色體分離、DNA修復和其他真核生物基本染色體過程。這些染色體過程的獨特需求推動了特定組蛋白變異體的出現。將這些變異體組蛋白整合到核小體中產生了染色質結構的巨大多樣性。
1. 什么是組蛋白變異體?
組蛋白變異體是核心組蛋白的非等位基因異構體,它們在基因組的戰略位置取代它們的經典組蛋白對應物。組蛋白變異體與核心組蛋白的不同之處在于,它們的差異范圍從氨基酸的輕微改變到結構上的重大差異。通常,經典組蛋白通常在基因組復制期間合成,其表達在S期達到高峰,而組蛋白變異體在整個細胞周期中表達,并以一種與復制無關的方式整合到染色質中。一些組蛋白變異體表現出組織特異性表達,限制在生殖細胞中。
通常,真核生物基因組中含有大量的復制相關(RC)組蛋白,而組蛋白變異體的數量有限,通常只有一個或幾個拷貝。雖然組蛋白變異體與它們的RC組蛋白對應物具有相似之處,但它們在其組蛋白折疊域(HFDs)方面顯示出明顯的特征,從而使它們在序列和功能方面有所區別。
此外,組蛋白變異體在其N-和C-末端尾部通常顯示出更大的差異。這些差異導致它們與不同的伴侶蛋白結合以進行沉積,并施加不同的翻譯后修飾。因此,這些變化賦予了組蛋白變異體通過改變染色質特性而具有的專門功能 [4,5]。
2. 組蛋白變異體的分類
所有組蛋白都可以存在為變異體,但復制相關(RC)組蛋白分化為組蛋白變異體的程度并不均勻。在哺乳動物中,有許多H2A變異體,較少的H3變異體,甚至更少的H2B和H4變異體。這種不均勻的分化可能與每種組蛋白在核小體中的位置及其在替代后修改核小體性質的潛力有關。
| 經典組蛋白 | 組蛋白變異體 | 功能 | |
|---|---|---|---|
| H2A | H2A.X | 分布在基因組各處,DNA修復,性染色體重塑 | |
| H2A.Z | H2A.Z.1 | 轉錄調控,異染色質組織,染色體分離,基因組穩定性,細胞增殖 | |
| H2A.Z.2 | 轉錄調控 | ||
| H2A.Bbd | 限制于脊椎動物,不包括非活性X染色體,轉錄激活 | ||
| macroH2A1.2 | 限制于脊椎動物,位于雌性哺乳動物細胞中非活性X染色體,轉錄抑制和高階染色質緊縮 | ||
| H2A.J | 與細胞增殖和炎癥相關的基因表達 | ||
| H2A.B | 核小體解穩定,活躍轉錄,DNA修復,DNA復制,剪接,精子發生 | ||
| mouse H2A.L (H2A.L.2) | 精子發生,H2A.L2顯示的組蛋白到蛋白轉變 | ||
| H2A.P | 睪丸特異性 | ||
| H2A.W | 植物特異性;保護比大多數核小體的147個堿基對多出的10-15個堿基對的連接區DNA | ||
| H2B | H2B.E | 小鼠氣味感受神經元 | |
| H2B.W | 睪丸特異性 | ||
| TH2B | 睪丸特異性,組蛋白到蛋白轉變 | ||
| H2B.3 | 在成熟葉片中富集,以及含有H3.3和/或H2A.Z的核小體 | ||
| H2B.8 | 在干燥的種子中富集 | ||
| H3 | H3.1 | 經典型,復制依賴性 | |
| H3.2 | 經典型,復制依賴性 | ||
| H3.3 | 與細胞周期無關,轉錄激活,端粒,著絲粒 | ||
| H3T (H3.4) | 睪丸特異性,在精母細胞和精子細胞中表達,在精子發生過程中的染色質轉變動態中起作用 | ||
| mouse H3t | 精子發生 | ||
| H3.5 | 位于人類染色體12的12p11.21區域,主要在生殖階段VI-X的精原細胞和初級精子細胞中表達,精子發生,轉錄調控, 在各種組織中弱表達 | ||
| H3.6 | 在各種組織中弱表達 | ||
| H3.7 | 在各種組織中弱表達 | ||
| H3.8 | 在各種組織中弱表達 | ||
| H3.X | 位于人類染色體5的5p15.1區域,轉錄調控,調節細胞周期 | ||
| H3.Y | 位于人類染色體5的5p15.1區域,轉錄調控,調節細胞周期 | ||
| CENP-A (CenH3) | 著絲粒特異性,有助于有絲分裂期間著絲粒組裝和染色體分離 | ||
| H3mm7 | 小鼠H3.3亞變體,表達在骨骼肌衛星細胞中 | ||
| H4 | H4-G | rDNA轉錄的上調 | |
| H1 | H1.1 | - | |
| H1.2 | 在小鼠胚胎干細胞中,H1.2和H1.3富集在H3K9me3標記的異染色質域上,通過阻滯RNAPII的轉錄,促使外顯子的包含 | ||
| H1.3 | |||
| H1.4 | |||
| H1.5 | 在人肺成纖維細胞中,H1.5富集在比核小體更短的外顯子的剪接位點上,通過阻滯RNAPII的活動促使它們被包含 | ||
| H1.6 | 睪丸特異性 | ||
| H1T2 | - | ||
| H1oo | - | ||
| HILS1 | - | ||
| H1X | 富集在RNAPII富集的區域;對星形膠質瘤不良預后的預示因素 | ||
| H1.0 | 在核小體相關域富集;在腫瘤細胞中通常呈不均勻表達,H1.0水平與腫瘤細胞分化和患者生存率相關 | ||
表1. 哺乳動物組蛋白變異體
表格信息來源: https://www.sciencedirect.com/science/article/pii/S0022283620305866和https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8015243/
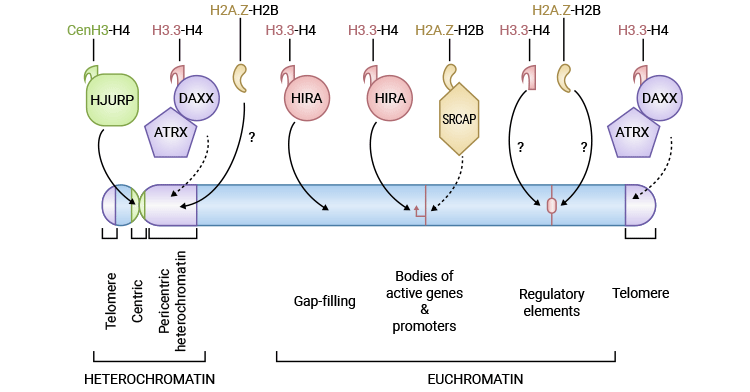
圖1. 組蛋白變體的特性
3. 組蛋白變體的功能
組蛋白變體通過影響組蛋白后轉錄修飾和組蛋白變體特異性結合伙伴的招募,影響核小體穩定性和高階染色質組織 [3,6]。組蛋白變體在幾乎所有DNA模板過程中發揮關鍵作用,例如DNA修復,染色體分離,轉錄調控,基因組穩定性,表觀遺傳,甚至組織特異性功能 [2,3]。
例如,著絲粒特異性變體著絲粒蛋白A(CENP-A)作為染色體著絲粒區的表觀遺傳標志,而H3.3與染色質動態和核小體周轉密切相關。此外,許多不同的H2A變體參與DNA修復,基因調控和其他關鍵過程,從而影響DNA組織。因此,組蛋白變體表達和沉積的失調可能導致發育綜合癥和癌癥。
4. 組蛋白變體與疾病
由于組蛋白變體具有普遍表達和多效性效應,它們與正常生理和許多疾病,包括發育性疾病和癌癥有關,這并不奇怪。例如,由于組蛋白變體可能是后期分化組織(如大腦)中新組蛋白的唯一來源,組蛋白變體可能在大腦相關疾病(如神經發育和神經退行性疾病)中發揮重要作用。實際上,一些研究表明,H3.3和H2A.Z變體對大腦功能至關重要 [7-9]。
此外,鑒于許多組蛋白變體是睪丸特異性的,替代或修改睪丸特異性組蛋白變體可能導致男性不育 [10]。
眾所周知,許多癌癥和發育性綜合癥攜帶編碼染色質相關蛋白,包括組蛋白變體及其沉積特異性伙伴的基因的復發性突變。研究表明,H3.3的突變在兒童神經膠質瘤和軟骨母細胞瘤中頻繁發現 [11, 12]。H2A.Z被證明具有致癌功能,而MacroH2A和H2A.X則抑制腫瘤發生。H2A.Z的上調在前列腺和膀胱癌中發現 [13, 14]。隨著黑色素瘤、膀胱癌和肛門腫瘤的疾病進展,MacroH2A的表達下降 [15, 16]。
參考文獻:
[1] S.G. Franklin, A. Zweidler. Non-allelic variants of histones 2a, 2b and 3 in mammals [J]. Nature, 266 (1977), pp. 273-275.
[2] Talbert PB, Henikoff S. 2010. Histone variants--ancient wrap artists of the epigenome [J]. Nat Rev Mol Cell Biol 11:264–275.
[3] Martire S, Banaszynski LA. 2020. The roles of histone variants in fine-tuning chromatin organization and function [J]. Nat Rev Mol Cell Biol 21:522–541.
[4] Henikoff S, Smith MM. 2015. Histone variants and epigenetics [J]. Cold Spring Harb Perspect Biol 7:a019364.
[5] Talbert PB, Henikoff S. 2017. Histone variants on the move: substrates for chromatin dynamics [J]. Nat Rev Mol Cell Biol 18:115–126.
[6] Buschbeck M., Hake S.B. Variants of core histones and their roles in cell fate decisions, development and cancer [J]. Nat. Rev. Mol. Cell Biol. 2017;18:299–314.
[7] Xia W & Jiao J Histone variant H3.3 orchestrates neural stem cell differentiation in the developing brain [J]. Cell Death Differ. 24, 1548–1563 (2017).
[8] Stefanelli G et al. Learning and age-related changes in genome-wide H2A.Z binding in the mouse hippocampus [J]. Cell Rep. 22, 1124–1131 (2018).
[9] Zovkic IB, Paulukaitis BS, Day JJ, Etikala DM & David Sweatt J Histone H2A.Z subunit exchange controls consolidation of recent and remote memory [J]. Nature 515, 582–586 (2014).
[10] Govin J et al. Pericentric heterochromatin reprogramming by new histone variants during mouse spermiogenesis [J]. J. Cell Biol 176, 283–294 (2007).
[11] Schwartzentruber J et al. Driver mutations in histone H3.3 and chromatin remodelling genes in paediatric glioblastoma [J]. Nature 482, 226–231 (2012).
[12] Behjati S et al. Distinct H3F3A and H3F3B driver mutations define chondroblastoma and giant cell tumor of bone [J]. Nat. Genet 45, 1479–1482 (2013).
[13] Dryhurst D., Ausió J. Histone H2A.Z deregulation in prostate cancer. Cause or effect [J]? Cancer Metastastis Rev. 2014;33:429–439.
[14] Kim K., Punj V., et al. Gene dysregulation by histone variant H2A.Z in bladder cancer [J]. Epigenetics Chromatin. 2013;6:1–13.
[15] Kapoor A., Goldberg M.S., et al. The histone variant macroH2A suppresses melanoma progression through regulation of CDK8 [J]. Nature. 2010;468:1105–1109.
[16] Hu W.-H., Miyai K., et al. Loss of histone variant macroH2A2 expression associates with progression of anal neoplasm [J]. J. Clin. Pathol. 2016;69:627–631.





