NRP1增強SARS-CoV-2感染能力的新機制
日期:2023-12-11 13:08:34
SARS-CoV-2感染的爆發引發了與COVID-19相關的全球大流行。隨著對病毒侵入機制的研究深入進行,一些潛在的機制已經被揭示出來。早在二月初,SARS-CoV-2侵入人體細胞的關鍵被確定:SARS-CoV-2-S與細胞表面的ACE2結合(更多信息);二月中旬,人體細胞中的TMPRSS2可以裂解ACE2和刺突蛋白的細胞表面蛋白酶,在病毒侵入過程中發揮了“協助”作用;三月,發現了第二條入侵途徑:細胞表面的CD147。
最近,在2020年10月20日,兩篇論文同時發表在《Science》雜志上。兩個科研團隊發現除了眾所周知的ACE2受體外,寄主細胞上另一個受體稱為 Neuropilin-1(NRP1)也會與SARS-CoV-2的S蛋白相互作用 [1] [2]。這一突破性的發現不僅有助于我們理解為何SARS-CoV-2比SARS病毒更具傳染性并引發更復雜的癥狀,還指出了新的潛在靶點,對于開發抗SARS-CoV-2藥物具有重要意義。
那么 Neuropilin-1是什么?SARS-CoV-2如何通過 Neuropilin-1侵入人體?您可以在以下內容中找到這些信息。
1. Neuropilin-1的定義
Neuropilin-1,又稱為血管內皮細胞生長因子165受體(VEGF165R)、CD304,屬于神經粘蛋白家族。神經粘蛋白(NRPs)最早由Takagi等人在爪蟾的神經組織冰凍切片中通過免疫熒光染色首次發現,并確認其為一種非酪氨酸激酶跨膜糖蛋白。NRPs家族中首次發現并引起關注的是NRP-1。
NRP-1分為兩種形式,膜型 Neuropilin-1(mNRP-1)和可溶性 Neuropilin-1(sNRP-1)。眾所周知,NRP-1通常指的是mNRP-1。如圖1所示,mNRP-1具有較大的N末端細胞外區域,一個短的跨膜區域,以及一個小的胞內細胞質區域,作為I型單通道跨膜蛋白[3]。
細胞外區域由三個部分組成:一個N末端的互補結合CUB(a1/a2)結構域,b1/b2結構域和meprin或MAM(c)結構域[4]。a1/a2結構域(或CUB結構域)在補體系統中介導蛋白質的相互作用。b1/b2結構域與凝血因子V和VIII的C1/C2結構域具有同源性。c結構域參與受體的二聚化。c結構域對于NRP-1的共受體活性至關重要。
短的跨膜螺旋結構域包含一個保守的GXXXG重復序列,據報道它能夠介導NRP-1的二聚化潛力,與c結構域一起。NRP-1的小的胞內片段缺乏酪氨酸激酶活性,但具有與PDZ蛋白相互作用的C末端SEA(Ser-Glu-Ala-COOH)基序,促進細胞內信號轉導。
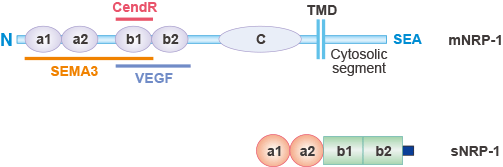
圖1. Neuropilin-1 (NRP1)的結構
*該圖片來源于Current Pharmaceutical Design的出版物 [3]
sNRP-1是一種相對分子質量約為9x104的蛋白,包含a1/a2和b1/b2結構域,但缺乏c結構域、跨膜區和胞內區。它是由NRP-1基因選擇性剪接產生的。它可以分泌到細胞外,進入血液循環,結合并抑制VEGF,從而負調控這些途徑介導的功能[5][6]。然而,sNRP-1的作用可能比簡單的誘騙作用更為復雜,sNRP-1的全部功能尚未完全了解[7]。
2. Neuropilin-1的功能
神經粘蛋白(NRPs)是一類單通道、非酪氨酸激酶表面糖蛋白,廣泛存在于所有脊椎動物中,種類間高度保守。已知存在兩種同源的NRP亞型,即NRP1和NRP2。這兩種NRPs最初被發現是神經元粘附分子。后來,它們被發現與血管生物學有關。正常的胚胎血管發育需要NRP1和NRP2參與小淋巴管和毛細血管的形成。過去10年的研究表明,NRP是一種多功能蛋白質,除神經和血管發育外,還參與多種生物過程。其中,NRP1在免疫和腫瘤發生中發揮關鍵作用。
Neuropilin-1之所以成為多功能信號傳導跨膜蛋白,是因為神經粘蛋白1作為一種共受體與多種細胞外配體相互作用,包括Sema3(與NRP-1的a1/a2和部分b1/b2結構域結合)、生長因子VEGF-A/B/D/E、胎盤生長因子2、HGF、TGF-B1、bFGF和PDGF(與NRP-1受體的b1/b2結構域結合)以及膜受體(如絲狀蛋白、VEGF受體、PDGF受體和TGF受體,與NRP-1受體的b1/b2和c結構域結合)[8] [9]。此外,NRP-1還可以與其他跨膜蛋白相互作用,如整合素αvβ3和β1,從而激活一系列信號傳遞途徑。
3. Neuropilin-1與SARS-CoV-2入侵的關系
與引起普通感冒和輕微呼吸道癥狀的其他冠狀病毒不同,引起COVID-19的SARS-CoV-2具有高度的傳染性和傳播性,并且臨床上COVID-19的非典型癥狀還包括味覺和嗅覺障礙、中風等。但是,先前的研究結果(ACE2、CD147和TMPRSS2-S蛋白)難以解釋為什么SARS-CoV-2容易感染呼吸系統以外的器官,比如大腦和心臟?這個問題促使科學家進一步研究SARS-CoV-2。
與SARS病毒不同,SARS-CoV-2的S蛋白在S1/S2界面上呈現多堿性序列基序RRAR。這個基序提供了一個酶切位點,叫做furin(一種宿主前蛋白酶)[10] [11]。因此,人體內的furin可以將S蛋白切割成兩個亞單位,S1和S2。當研究人員觀察SARS-CoV-2 S1序列時,他們發現該序列的一個片段與人體蛋白中與NRP1相互作用的蛋白序列非常相似。基于這一發現,研究人員提出了一個簡單的假設:SARS-CoV-2 S1能否與NRP1結合,進一步幫助病毒感染人體細胞?然后,兩個研究團隊通過不同的方法證實了這一假設。
德國慕尼黑工業大學的一個研究團隊使用體外培養的細胞證明,在ACE2存在的情況下,NRP-1可以促進病毒感染。一個具有改變酶切位點的SARS-CoV-2突變體不依賴于NRP1的感染能力。此外,對人類COVID-19尸檢的病理分析顯示,SARS-CoV-2感染的細胞,包括面對鼻腔的嗅神經元,NRP1呈陽性。Simons教授解釋說,如果將ACE2視為細胞的入口,那么NRP1可能是將病毒帶到細胞門口的因素。
英國布里斯托大學的另一個研究團隊也通過一系列結構和生化方法確定了SARS-CoV-2 S1與NRP1直接結合。更重要的是,這兩個研究團隊在體外培養的人類細胞系中使用針對NRP1的單克隆抗體或選擇性NRP1抑制劑阻止了S蛋白與NRP1之間的相互作用。實驗結果顯示,SARS-CoV-2的感染能力顯著降低。
4. Neuropilin-1與癌癥
神經粘蛋白1(NRP1)在哺乳動物體內的神經元、血管、免疫細胞和許多其他細胞類型中都有表達。它結合了一系列結構和功能多樣的細胞外配體,以調節器官的發育和功能。近年來,已開發了幾種類型的小鼠基因敲除模型,為研究NRP1功能提供了有用的工具,并設計了大量以癌癥治療為目的的靶向NRP1的治療藥物,其中大部分是為了探索癌癥治療而設計的[12]。
在小鼠中,NRP1選擇性地在胸腺源性T調節細胞上表達,并極大地增強了免疫抑制功能。在人體中,NRP1在漿細胞樣樹突細胞(pDCs)上表達,有助于啟動免疫反應,并在某些調節性T細胞(Tregs)的亞群上表達[13] [14]。初步研究表明,T細胞上的NRP1表達使其具有增強的免疫抑制活性[15]。已在炎癥微環境中分離出的激活T細胞和Tregs上鑒定出NRP1的表達。這些研究表明,NRP1可能是一種新的T細胞激活標記。在臨床方面,NRP1可能增強Treg腫瘤浸潤,NRP1+ Tregs的減少與成功的化療相關,提示NRP1在癌癥病理中具有特定作用。作為一個治療靶點,NRP1可以同時靶向NRP1表達的腫瘤血管、NRP1+ Tregs和pDCs [16] [17] [18]。
目前已知,靶向NRP1的抗腫瘤治療可以通過不同的工具實現,包括sNRP-1、單克隆抗體、肽和肽類似物、小干擾RNA或微RNA以及細胞穿透肽[19] [20]。NRP1代表了癌癥治療的一個有前途的新靶點。目前,一種名為Vesencumab的單克隆抗體已由Genentech Inc.開發并進行了臨床試驗。
參考文獻:
[1] James Daly et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection [J]. Science. 2020.
[2] LUDOVICO CANTUTI-CASTELVETRI et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity [J]. Science. 2020.
[3] Lukui Chen, Guilong Zhang, et al. Neuropilin-1 (NRP-1) and Magnetic Nanoparticles, a Potential Combination for Diagnosis and Therapy of Gliomas [J]. Curr. Pharm. Des. 2015, 21(37): 5434-5449.
[4] Gu CH, Limberg BJ, et al. Characterization of neuropilin-1 structural features that confer binding to semaphorin 3A and vascular endothelial growth factor 165 [J]. J Biol Chem 2002, 277(20): 18069-76.
[5] Cackowski FC, Xu L, et al. Identification of two novel alternatively spliced neuropilin-1 isoforms [J]. Genomics 2004; 84(1): 82-94.
[6] Uniewicz KA, Cross MJ, et al. Exogenous recombinant dimeric neuropilin-1 is sufficient to drive angiogenesis [J]. J Biol Chem 2011; 286(1): 12-23.
[7] Belal Chaudhary, Yazan S. Khaled, et al. Neuropilin 1: function and therapeutic potential in cancer [J]. Cancer Immunol. Immunother. 2013.
[8] Graziani G, Lacal PM. Neuropilin-1 as Therapeutic Target for Malignant Melanoma [J]. Front Oncol. 2015, 3;5:125.
[9] Yaqoob U, Cao S, et al. Neuropilin-1 stimulates tumor growth by increasing fibronection fibril assembly in the tumor microenvironment [J]. Cancer Res. 2012, 72(16): 4047-4059.
[10] D. Wrapp, N. Wang, K. S. Corbett, et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation [J]. Science. 2020, 367, 1260–1263.
[11] M. Hoffmann, H. Kleine-Weber, et al. A multibasic cleavage site in the spike protein of SARS-CoV-2 is essential for infection of human lung cells [J]. Mol. Cell. 2020, 78, 779–784.e5.
[12] Claudio Raimondi, James T. Brash, et al. NRP1 function and targeting in neurovascular development and eye disease [J]. Progress in Retinal and Eye Research. 2016, 1-20.
[13] Romeo PH, Lemarchandel V, et al. Neuropilin-1 in the immune system [J]. Adv Exp Med Biol. 2002, 515:49–54.
[14] Tordjman R, Lepelletier Y, et al. A neuronal receptor, neuropilin-1, is essential for the initiation of the primary immune response. Nat Immunol. 2002, 3(5):477–482.
[15] Yuan Q, Hong S, et al. CD4(+)CD25(?)Nrp1(+) T cells synergize with rapamycin to prevent murine cardiac allorejection in immunocompetent recipients [J]. PLoS ONE. 2013, 8(4):e61151.
[16] Chaudhary B, Elkord E. Novel expression of Neuropilin 1 on human tumor-infiltrating lymphocytes in colorectal cancer liver metastases [J]. Expert Opin Ther Targets. 2015, 19 (2): 147-161.
[17] Bourbie-Vaudaine S, Blanchard N, et al. Dendritic cells can turn CD4+ T lymphocytes into vascular endothelial growth factor-carrying cells by intercellular neuropilin-1 transfer [J]. J Immunol. 2006, 177(3):1460–1469.
[18] Battaglia A, Buzzonetti A, et al. Neuropilin-1 expression identifies a subset of regulatory T cells in human lymph nodes that is modulated by preoperative chemoradiation therapy in cervical cancer [J]. Immunology. 2008, 123(1):129–138.
[19] Teesalu T, Sugahara KN, et al. Tumor-penetrating peptides [J]. Front Oncol. 2013, 3:216.
[20] Teesalu T, Sugahara KN, et al. C-end rule peptides mediate neuropilin-1-dependent cell, vascular, and tissue penetration [J]. Proc Natl Acad Sci USA. 2009, 106(38):16157–16162.









