SARS-CoV-2感染會發生抗體依賴性增強嗎?
日期:2023-12-11 13:46:42
在1964年,Hawkes等人證明了病毒在病毒特異性抗血清(具體是IgG抗體)的亞中和濃度下,包括蟲媒病毒、穆雷河谷腦炎病毒、西尼羅河病毒(WNV)、日本腦炎病毒(JEV)和格他病毒等病毒的增強感染能力 [1]。這是首次描述的“抗體依賴性感染增強”(ADE)。ADE最早是由斯科特·哈爾斯特德(Scott Halstead)在1973年首次在登革病毒(DENV)感染中描述的 [2] [3]。此后還發現其他病毒也表現出ADE,包括流感A病毒、科薩奇病毒B、呼吸道合胞病毒、埃博拉病毒和人類免疫缺陷病毒(HIV)。
在本文中,我們將介紹ADE的定義、ADE的機制、ADE研究的意義、ADE是否在SARS-CoV-2感染中發生以及減少ADE風險的解決方案。
1. ADE的定義
病毒感染都是通過與細胞表面黏附開始的。黏附是通過病毒表面蛋白與目標細胞上特定受體之間的相互作用來完成的。病毒特異性抗體通常可以阻止這一步驟,或者“中和”病毒,使病毒失去感染細胞的能力 [4]。然而,在某些情況下,抗體并不是在病毒感染過程中防止病毒入侵人體細胞,而是輔助病毒進入目標細胞,增強病毒在體內的復制,并提高感染率,引發嚴重的病理反應。這種現象被稱為病毒感染的抗體依賴性增強效應(ADE)。
發生ADE的病毒多半在巨噬細胞內增殖,并易導致宿主持續感染。常規疫苗通常難以預防和治療這類病毒性疾病 [5],在某些情況下已被證明會增加疫苗接種者感染的風險。
2. 抗體依賴性增強的機制
ADE有兩種機制,這兩種機制都發生在非中和抗體或亞中和濃度的抗體與病毒抗原結合而未防止或清除感染的情況下。抗體-病毒復合物通過IgG抗體的Fc部分與Fcγ受體(FcγR)或補體受體之間的相互作用附著在細胞上,并增加了病毒的內吞作用,導致病毒復制增強和疾病嚴重性加劇。一種是FcγR介導的ADE(FcγR-ADE),涉及病毒、抗體和細胞表面的FcγR;另一種是補體介導的ADE(C-ADE),與病毒、抗體、補體和補體受體有關。
2.1 FcγR介導的ADE
在1977年,斯科特·哈爾斯特德(Scott Halstead)等人發現,一些感染了一種登革病毒血清型的人,可能在與異源性登革病毒血清型的二次感染時發展出嚴重疾病,并將登革病的臨床嚴重程度與ADE聯系起來 [6] [7]。這是關于片段結晶區受體(FcR)介導的ADE的首次報道。FcR依賴性ADE被認為是ADE的最常見機制。ADE是在抗體結合的病毒通過Fc-FcγR相互作用與攜帶FcγR的細胞(包括巨噬細胞、單核細胞、樹突狀細胞、中性粒細胞、白細胞和粒細胞)結合時開始的,通過FcγR介導的內吞作用或吞噬作用促進抗體包被的病毒進入細胞 [8]。FcR介導的ADE在許多病毒中都有發現,包括人類免疫缺陷病毒(HIV)、阿留申貂細小病毒、呼吸道合胞病毒、西尼羅河病毒和流感A病毒。
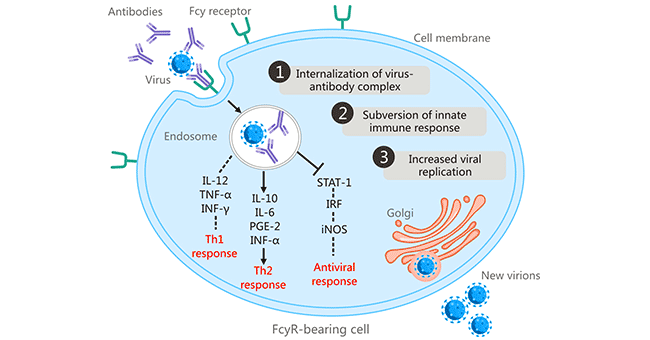
圖1. fcr介導的抗體依賴性增強
圖片來自https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6290032/
2.2 補體介導的ADE
在補體介導的ADE中,補體包被蛋白C1q結合到與病毒結合的抗體的Fc區域上 [9],這改變了C1s C1r復合物的構象,激活了C1s C1r復合物。激活的C1s裂解補體C2和C4,所產生的裂解產物形成了C3轉換酶(C4bC2a)。C3轉換酶將C3裂解為C3a和C3b。高反應性的C3b與病毒和攜帶CR的細胞上的CR結合,通過吞噬作用導致病毒進入細胞,從而增強病毒復制并加重疾病嚴重性 [10]。C1q受體不僅分布在炎癥性單核/巨噬細胞上,還分布在其他不同類型的細胞上,包括B細胞、成纖維細胞、中性粒細胞和內皮細胞。這種機制是西尼羅河病毒和HIV中ADE的基礎。
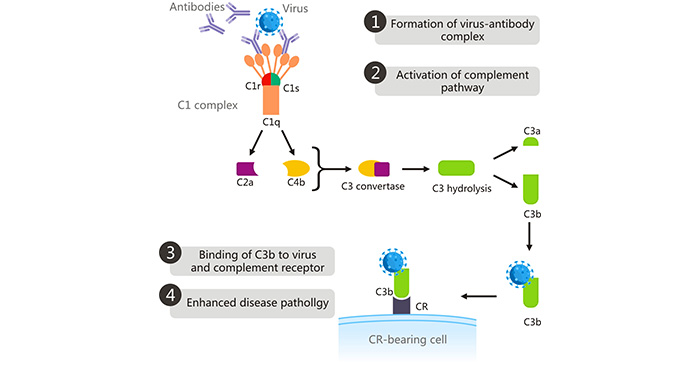
圖2. 補體介導的ADE
圖片來自https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6290032/
除了常見的FcR介導的ADE外,埃博拉病毒(EBOV)已被發現可利用補體成分C1q引發感染的ADE。這種機制通過交聯病毒-抗體-C1q復合物與細胞表面C1q受體結合來介導,從而增強病毒進入細胞。在C1q依賴的ADE中,兩個或更多的IgG單體抗體從病毒-抗體復合物突出,與C1q結合形成病毒-抗體-C1q復合物 [11]。病毒-抗體-C1q復合物與靶細胞上的C1q受體結合,觸發Wnt/β-catenin、PI3K和一些酪氨酸激酶等信號通路,促進通過內吞作用使抗體包被的病毒進入靶細胞。在某些情況下,C1q在HIV感染背景下直接結合到病毒的外膜糖蛋白gp41上。
3. ADE研究的意義
不同病毒感染的ADE效應與目標細胞類型、目標細胞的分化和成熟程度以及分化程度有關。此外,研究發現ADE效應是否在病毒感染中發生也與病毒感染和被動免疫之間的時間間隔有關。如果特異性抗體與病毒同時注射動物,大多數情況下可以發揮協調的保護作用。在感染病毒后注射抗體,甚至中和抗體也可能增強病毒的毒力。
更好地了解ADE的各種機制在確定與病毒相關的抗原決定因素方面起著關鍵作用。針對并修改這些抗原決定因素將有助于開發更安全、更有效的疫苗。更重要的是,ADE效應的性質是機體免疫調節效應的一部分。清晰的機制可能為發展針對具有明確病原體感染的傳染病的新藥提供機會和方向。
然而,我們仍然可以理解,ADE對于未來免疫治療,尤其是針對病原體感染引起的傳染病的疫苗開發具有深遠和啟發性的意義。在未來,醫學領域仍將聚焦于這一熱點話題,并為研究和開發新藥提供更多信息。
4. SARS-CoV-2感染或COVID-19疫苗接種是否存在抗體依賴增強跡象?
由于COVID-19疫苗引起的ADE效應一直是公眾關注的焦點。最新的科學研究發現,在體外,能中和病毒和增強感染的受體結合區(RBD)抗體或感染增強的N端區(NTD)抗體可以在非人靈長類動物和小鼠中保護免受SARS-CoV-2感染 [17]。與引發病毒載量和炎癥增加等顯著疾病惡化不同,這些抗體展現了部分保護效果。文章的作者認為,SARS-CoV-2在巨噬細胞中無法有效復制(Fc介導的ADE抗體主要進入巨噬細胞),而Fc介導效應帶來的保護效果可能有助于這個實驗結果。總之,現有的證據不足以支持SARS-CoV-2感染或COVID-19疫苗接種會引發明顯的ADE效應。
雖然在MERS-CoV和SARS-CoV感染中已經顯示出ADE,但迄今尚未有關于SARS-CoV-2感染或COVID-19疫苗接種引發ADE的確認報告。在人類研究中,之前感染過冠狀病毒的人在感染不同類型的冠狀病毒后并未出現增強的疾病。在COVID-19疫苗開發過程中,科學家采取了幾種方法來避免ADE。例如,在早期疫苗設計中選擇最不可能引發ADE的SARS-CoV-2蛋白質作為抗體靶點。對動物進行疫苗接種以觀察是否發生ADE。評估臨床試驗中人類和患者的情況。在實際COVID-19疫苗數據中尋找案例。
5. 減少ADE風險的解決方案
ADE已在40多種病毒中被確認。它對病毒感染的抗體治療和疫苗開發產生了負面影響。抗體疫苗和療法的一個潛在障礙是通過ADE加劇疾病的嚴重性的風險。已經證明ADE參與了許多病毒感染的疾病加重,包括寨卡病毒、埃博拉病毒 [12]、呼吸道合胞病毒(RSV)、麻疹、SARS-CoV和MERS-CoV。呼吸道感染中的ADE被包含在更廣泛的類別中,稱為增強型呼吸道疾病(ERD),還與細胞因子級聯和細胞介導的免疫病理學等非抗體機制有關。
疫苗是針對健康個體的常規醫學干預措施,因此在開發新疫苗時必須考慮ADE現象,以確保疫苗具有保護作用且不會對接種者造成傷害。疫苗開發者尋求方法來降低ADE的可能性。
在冠狀病毒感染中降低ADE風險的方法有幾種。第一種方法是誘導或提供高劑量的強效中和抗體。Wan等人指出,高劑量的抗體可以在MERS-CoV中抑制ADE,而不會干擾其抗病毒能力 [13]。第二種方法是在早期疫苗設計中選擇最不可能引發ADE的病毒蛋白作為抗體靶點。對于SARS-CoV-2來說,選擇刺突(S)蛋白的受體結合區(RBD)內的特定表位作為中和抗體反應的靶點。令人鼓舞的是,一些早期臨床試驗展示了強烈的中和抗體反應和強大的Th1反應,而不是與免疫病理相關的Th2反應 [14]。利用一些抑制劑還有助于降低ADE的風險。蛋白酶抑制劑和Fc抑制劑已被證明在MERS-CoV和SARS-CoV中發揮了抑制作用 [13][15][16]。此外,通過修改抗體Fc區域上的FcγR結合位點來降低登革病毒的ADE風險。
參考文獻:
[1] Hawkes R. A. Enhancement of the infectivity of arboviruses by specific antisera produced in domestic fowls [J]. Aust. J. Exp. Biol. Med. Sci. 1964, 42, 465–482.
[2] Halstead, S. B., Chow, J. & Marchette, N. J. Immunologic enhancement of dengue virus replication [J]. Nat. New Biol. 243, 24–25 (1973).
[3] Halstead, S. B., Shotwell, H. & Casals, J. Studies on the pathogenesis of dengue infection in monkeys [J]. II. Clinical laboratory responses to heterologous infection. J. Infect. Dis. 128, 15–22 (1973).
[4] Dutta, A, Huang, CT, et al. (2016). Sterilizing immunity to influenza virus infection requires local antigen-specific T cell response in the lungs [J]. Scientific Reports. 6: 32973.
[5] Wang Q., Zhang L., et al. Immunodominant SARS coronavirus epitopes in humans elicited both enhancing and neutralizing effects on infection in non-human primates [J]. ACS Infect Dis. 2016; 2: 361-376.
[6] Halstead SB, O'Rourke EJ. Dengue viruses and mononuclear phagocytes. I. Infection enhancement by non-neutralizing antibody [J]. J Exp Med 1977;146(1):201–17.
[7] Halstead SB. Immune enhancement of viral infection [J]. Prog. Allergy31, 301–364 (1982).
[8] Taylor A., Foo S. S., et al. Fc receptors in antibody-dependent enhancement of viral infections [J]. Immunol. Rev. 2015, 268, 340–364.
[9] Diebolder CA, Beurskens FJ, et al. Complement is activated by IgG hexamers assembled at the cell surface [J]. Science. (2014) 343:1260– 3.
[10] Takada A., Kawaoka Y. Antibody-dependent enhancement of viral infection: molecular mechanisms and in vivo implications [J]. Rev. Med. Virol. 2003, 13, 387–398. (complement dependent ADE)
[11] von Kietzell K., Pozzuto T., et al. Antibody-mediated enhancement of parvovirus B19 uptake into endothelial cells mediated by a receptor for complement factor C1q [J]. J Virol. 2014; 88: 8102-8115.
[12] Takada, A., Feldmann, H., et al. Antibody-dependent enhancement of Ebola virus infection [J]. J. Virol. 77, 7539–7544 (2003).
[13] Y. Wan, J. Shang, et al. Molecular mechanism for antibody-dependent enhancement of coronavirus entry [J]. J Virol, 94 (5) (2020).
[14] Anderson EJ, Rouphael NG, Widge AT, et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults [J]. N Engl J Med 2020; 383:2427-2438.
[15] L. Liu, Q. Wei, et al. Anti-spike IgG causes severe acute lung injury by skewing macrophage responses during acute SARS-CoV infection [J]. JCI Insight, 4 (4) (2019).
[16] Jieqi Wen, Yifan Cheng, et al. Antibody-dependent enhancement of coronavirus [J]. International Journal of Infectious Diseases, Volume 100, November 2020, Pages 483-489.
[17] Dapeng Li, Robert J Edwards, et al. The functions of SARS-CoV-2 neutralizing and infection-enhancing antibodies in vitro and in mice and nonhuman primates [J]. Version 2. bioRxiv. Preprint. 2021 Jan 2 [revised 2021 Feb 18].









