Oct4——胚胎干細胞多能性的守門人
日期:2023-11-01 14:36:00
Oct4(八聚體結合轉錄因子)之名來自于其與八聚體基序DNA結合的能力。它還被稱為Oct3、POU5F1、Oct-3/4、Otf3或NF-A3。它由POU5F1基因編碼,屬于POU轉錄因子家族的第5亞家族。Oct4是胚胎干細胞多能性的守門人,對于維持胚胎干細胞的多能性和自我更新起著極其重要的作用。Oct4還對誘導多能干細胞的產生至關重要。此外,Oct4在多種腫瘤中高度表達,這表明Oct4可能參與惡性腫瘤的發生和發展。
1. Oct4的組織特異性
Oct4在發育中的大腦中表達,高水平表達于皮層、嗅球、海馬和小腦等特定細胞層。此外,Oct4主要在胚胎干細胞、胚胎生殖細胞和胚胎/生殖細胞腫瘤中表達。雖然成年組織中Oct4的表達水平較低,但在各種成年干細胞中廣泛表達,主要包括:
● 來自乳腺、胰腺、胃、肝臟、腎臟和間充質組織的干細胞。
● 前體細胞,如外周血、乳腺上皮、子宮內膜和卵泡。
● 多能性前體細胞等。
體外,Oct4在未分化的胚胎干細胞(ES細胞)、胚胎癌細胞(EC細胞)和胚胎生殖細胞中具有高表達水平。隨著這些細胞的分化,Oct4的表達水平下調 [1]。
在人類中,Oct4 mRNA在從未受精的卵母細胞到未壓縮的囊胚階段的胚胎發育的各個階段均有表達 [2]。
2. 什么是Oct4?
2.什么是Oct4?
Oct4由Pou5fl基因編碼,位于人類染色體6p21.3上。Oct4是POU轉錄因子家族的成員。它具有多個轉錄起始位點,可以轉錄成不同的mRNA亞型,從而翻譯成多種蛋白質。
2.1 Oct4的亞型
人類Oct4基因可以通過選擇性剪接產生三個亞型:Oct4A、Oct4B和Oct4B1 [3] [4]。Oct4可以翻譯成四個蛋白亞型(Oct4A、oct4b-190、oct4b-265和oct4b-164)[5]。
2.1.1 Oct4A
Oct4A轉錄本包含外顯子1、2b、2d、3和4。外顯子1是Oct4A的特定部分。Oct4A由360個氨基酸組成,主要在未分化細胞的細胞核中表達。它是多能干細胞的特異性轉錄因子,與細胞的未分化特性相關。
2.1.2 Oct4B
Oct4B包含外顯子2a、2b、2d、3和4。與Oct4A相比,Oct4B沒有外顯子1,但具有特定的外顯子2a。
Oct4B由265個氨基酸組成,主要在各種非多能性細胞中表達,例如末分化的外周血單核細胞和膀胱腫瘤細胞。
Oct4B與Oct4A在POU結合區域和羧基端部分有相似的結構,但在氨基端的轉錄激活區域有所不同[6]。Oct4B的氨基端抑制了與DNA的結合,并抑制了Oct4激活因子的轉錄活性,從而失去了細胞多能性的調控功能。
2.1.3 Oct4B1
Oct4B1轉錄本與Oct4B非常相似,但包含額外的外顯子2c。
OctB1主要在人類胚胎干細胞和胚胎癌細胞(ECC)中表達,其表達在細胞分化時迅速下調。
OctB1與Oct4B在POU結合區域和羧基端部分相同,其余氨基酸序列不同。Oct4B1增強了細胞的抗凋亡潛能,調節了細胞的多能狀態。此外,Oct4B1也參與了腫瘤的進展。
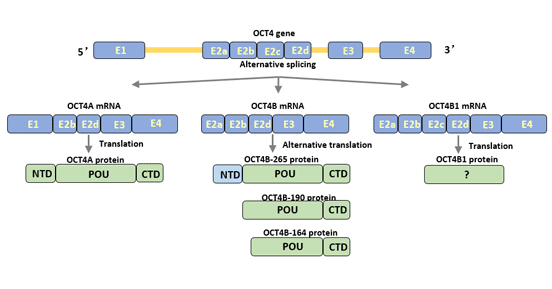
圖1. Oct4亞型的結構
2.2 Oct4基因的結構
Oct4亞型1是主要的轉錄亞型之一,含有5個外顯子和4個內含子。這個基因編碼的蛋白質包含324個氨基酸,長度約為16.40 kb,相對分子質量為18 kD。
Oct4由三個結構域組成:N-轉錄域、POU結合域和C-轉錄域。
N-端區域:這個區域是一個轉錄激活區域,富含脯氨酸,具有與DNA結合和激活轉錄的功能,有助于穩定生物分子結構,降低細胞酸性和調節細胞氧化還原潛能。
POU結合域:一個保守的DNA結合域,結合含有八聚體序列的DNA,以調控下游靶基因的轉錄。
POU結合域包含兩個亞基:N-端和C-端。N-端是POU家族(POUs)的獨特保守結構域,富含脯氨酸和酸性殘基。C-端是傳統的異源域(POUh),富含脯氨酸、絲氨酸和蘇氨酸殘基。POUs和POUh之間由一個連接肽相連。
C-端區域:富含絲氨酸/蘇氨酸,控制著不同細胞類型中Oct4A的轉活性。
2.3 Oct4偽基因
目前已經發現了六個Oct4偽基因,分別是Oct4pg1、Oct4pg2、Oct4pg3、Oct4pg4、Oct4pg5和Oct4pg6。這些偽基因與Oct4 mRNA高度同源,但不具有Oct4的功能。Oct4偽基因的存在可能會在實驗結果中引起假陽性,這給腫瘤和干細胞Oct4的研究帶來了一些困擾。
3. Oct4基因的調控
Oct4的上游調控元件包括近端增強子(PE)和遠端增強子(DE),在發育的不同階段具有不同的功能。
例如,近端增強子作用于外胚層干細胞,而遠端增強子則作用于內細胞團、原始生殖細胞和胚胎干細胞 [7]。許多因子通過作用于這兩個調控元件來調控Oct4的表達。例如,孤兒核受體超家族因子可以結合到PE并影響Oct4的表達。
此外,一些物質,如Paf1復合物,可以直接與Oct4啟動子相互作用,參與Oct4表達的激活。作為Oct4激活因子,它在維持胚胎干細胞Oct4的轉錄激活中起著重要作用。
Oct4也受到Sox2的調控。Sox2可以直接或間接調控Oct4的表達,從而影響胚胎干細胞的多能性。
Oct4蛋白的活性主要受到磷酸化或磺化的調控。
例如,通過蛋白激酶A和/或ERK MAPK在Oct4蛋白的POU結合域中高度保守的殘基Ser 229(小鼠)或Ser 236(人類)的磷酸化可以在空間上阻礙DNA結合和同源二聚體的組裝 [8]。
Oct4參與了幾個信號通路的調節,包括轉化生長因子(TGF)β通路、成纖維細胞生長因子(FGF)通路、磷脂酰肌醇-3-激酶(PI3K)通路、Wnt通路等。在心臟中胚層的特異分化過程中,TGF-β信號通路和經典Wnt信號通路共同誘導了Oct4的產生 [9]。
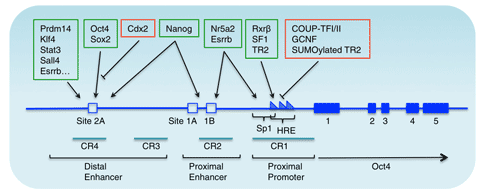
圖2. 小鼠Oct4基因的基因組結構及轉錄調控 [10]
4. Oct4的功能
Oct4通過與啟動子或增強子區域中的ATGCAAAT結合來調控靶基因的表達[11] [12]。這些靶基因主要包括YES1、FGF4、UTF1、ZFP206等與哺乳動物胚胎發育控制有關的基因。通過調控其下游靶基因,它參與了正常發育,尤其是在早期胚胎發育中。此外,Oct4的表達與干細胞的多能性有關。
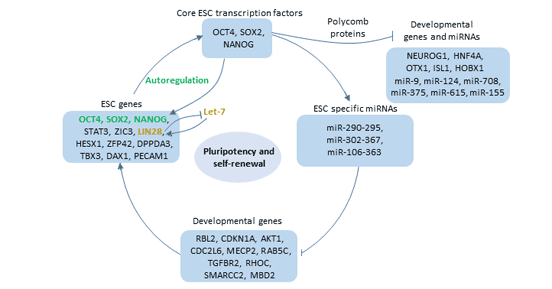
圖3. Oct4的功能
4.1 Oct4與胚胎發育
胚胎干細胞(ESCs)是一類具有自我更新能力和多向分化潛能的細胞,可以分化為身體各種類型的組織細胞。Oct4是ESCs中的一個重要轉錄因子,在維持ESCs的多能性和自我更新能力方面發揮著關鍵作用。
Oct4通過激活多能性基因并抑制分化基因的表達,使ESCs保持自我更新和未分化狀態。
Oct4在體內并不單獨起作用。Oct4與Nanog和Sox2一起構成了多能性調控網絡的核心,并被認為位于細胞全能性調控網絡的頂端。這三個轉錄因子結合到自身基因的啟動子上 [13],激活了自身基因的表達。
一方面,它們激活了與維持ES細胞自我更新有關的基因的轉錄。例如,Dppa4、Tdgf1、Oct4、Nanog和Lefty2都受到Oct4、Sox2和Nanog的正調控,并且在ES細胞中優先表達。
另一方面,Oct4、Sox2和Nanog共同占據與分化和發育有關的基因的啟動子,并促使它們失活 [14]。
Oct4作為轉錄調節因子在轉錄過程中具有雙重調節作用(轉錄抑制和轉錄激活),它不僅可以維持干細胞的未分化狀態,還可以控制它們向特定細胞類型的分化。
在胚胎發育過程中,Oct4的水平決定了干細胞的分化或去分化,ES細胞的命運由Oct4的表達精確調控。當Oct4過度表達時,胚胎干細胞分化為原始內胚層和中胚層細胞。當Oct4的表達水平較低時,ES細胞失去多能性,分化為滋養層細胞。只有在適當的表達水平下,胚胎干細胞才能保持其自我更新狀態。
研究發現,缺乏Oct4的胚胎只發育到囊胚階段,內胚層的細胞不是多能性的,只能分化為滋養層細胞。而囊胚在植入時就死亡了 [15]。
在成體干細胞中,Oct4維持多能性的調節機制與ESC類似。
4.2 延緩細胞老化
Oct4延緩細胞老化的能力對于老化研究具有重大意義。Oct4在所有生物體的發育中都很重要,并在細胞的整個生命周期中保持活性。它保護血管中的動脈粥樣硬化斑塊,這是心臟病和中風的根本原因。
4.3 Oct4在重編程中的作用
細胞狀態轉變通常被認為是從較不分化的狀態向較分化的狀態轉變,但研究表明,從體細胞向原始多能性的細胞狀態轉變也是可能的。這個過程是由于過表達Oct4、Sox2、Klf4和c-Myc等轉錄因子而誘導的。
其中,Oct4被認為是重編程中不可或缺的核心轉錄因子。Oct4調節細胞狀態轉變,如建立多能性和胚胎生殖細胞的分化。
通過將Oct4與Sox2、Klf4和c-Myc一起引入體細胞中,可以獲得誘導多能干細胞(iPSCs)。iPSCs是從不同類型和類型的體細胞中獲得的。
在上皮間質轉變過程中,Oct4可以抑制TGF beta R3和TGF beta 3,同時激活miR-200 miRNA家族,從而分別抑制上皮間質轉變(EMT)調節因子Snail和Zeb2,并最終促進間質向上皮轉變(MET)。
Oct4、Sox2和Klf4引導重編程過程。Oct4A等外源性重編程因子可以直接或間接地沉默體細胞基因并激活多能性基因。內源性Oct4A和其他多能性基因可以維持誘導多能干細胞的多能狀態。
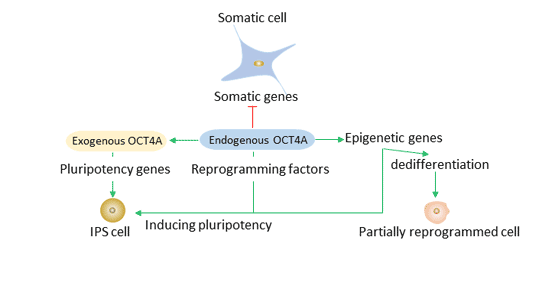
圖4. Oct4在重編程中的作用
4.4 Oct4與腫瘤發生性
Oct4在胚胎生殖細胞瘤和成人細胞腫瘤中高度表達,這表明Oct4可能參與了惡性腫瘤的發生和發展。
Nanog和Oct4在乳腺癌組織中高度表達 [16]。Oct4的表達與前列腺癌的分化程度有關 [17]。在口腔鱗狀細胞癌中,Nanog、Oct4和CD133的強表達與不良臨床預后有關。Oct4的表達是維持某些肺癌細胞干細胞特性的關鍵 [18]。
多項研究表明,Nanog和Oct4在多種不受限制、分化差、侵襲性固體腫瘤中高表達,而它們的下調可能抑制腫瘤發展 [19]。
Oct4的過表達可能導致腫瘤發生、腫瘤轉移以及化療和放療后的遠處復發 [20] [21]。
因此,通過抑制Oct4的表達可以降低腫瘤細胞的自我更新能力。
研究表明,Oct4可能參與維持腫瘤干細胞的生存,沉默Oct4有望成為針對腫瘤干細胞的新方法。Oct4對腫瘤的生長、轉移和復發有顯著影響。研究表明,Oct4在人類乳腺癌細胞系7(MeF7)細胞中高度表達,通過RNAi技術沉默Oct4可以促進腫瘤干細胞的凋亡。
參考文獻:
[1] Cauffman G, Van de Velde H, Liebaers I, et al. Oct-4 mRNA and protein expression during human preimplantation development [J]. Molecular human reproduction, 2004, 11(3): 173-181.
[2]Hansis C, Grifo J A, Krey L C. Oct-4 expression in inner cell mass and trophectoderm of human blastocysts [J]. Molecular human reproduction, 2000, 6(11): 999-1004.
[3]Takeda J, Seino S, Bell G I. Human Oct3 gene family: cDNA sequences, alternative splicing, gene organization, chromosomal location, and expression at low levels in adult tissues [J]. Nucleic acids research, 1992, 20(17): 4613-4620.
[4]Atlasi Y, Mowla S J, Ziaee S A M, et al. OCT4 spliced variants are differentially expressed in human pluripotent and nonpluripotent cells [J]. Stem cells, 2008, 26(12): 3068-3074.
[5]Wang X, Dai J. Concise review: isoforms of OCT4 contribute to the confusing diversity in stem cell biology [J]. Stem cells, 2010, 28(5): 885-893.
[6]Cauffman G, Liebaers I, Van Steirteghem A, et al. POU5F1 isoforms show different expression patterns in human embryonic stem cells and preimplantation embryos [J]. Stem cells, 2006, 24(12): 2685-2691.
[7]Shi G, Jin Y. Role of Oct4 in maintaining and regaining stem cell pluripotency [J]. Stem cell research & therapy, 2010, 1(5): 39.
[8]Brumbaugh J, Hou Z, Russell J D, et al. Phosphorylation regulates human OCT4 [J]. Proceedings of the National Academy of Sciences, 2012, 109(19): 7162-7168.
[9]Li Y, Yu W, Cooney A J, et al. Brief report: Oct4 and canonical Wnt signaling regulate the cardiac lineage factor Mesp1 through a Tcf/Lef-Oct4 composite element [J]. Stem cells, 2013, 31(6): 1213-1217.
[10] Wu G, Sch?ler H R. Role of Oct4 in the early embryo development [J]. Cell regeneration, 2014, 3(1): 7.
[11] Sch?ler, H.R, Balling R, Hatzopoulos A K, et al. Octamer binding proteins confer transcriptional activity in early mouse embryogenesis [J]. The EMBO Journal, 1989, 8(9):2551-2557.
[12]YOUNGILYEOM, Ha H, Balling R, et al. Structure, expression and chromosomal location of the Oct-4 gene [J]. Mechanisms of Development, 1991, 35(3): 171-179.
[13]Niwa H. How is pluripotency determined and maintained? [J]. Development, 2007, 134(4): 635-646.
[14]Christophersen N S, Helin K. Epigenetic control of embryonic stem cell fate [J]. Journal of Experimental Medicine, 2010, 207(11): 2287-2295.
[15]Chambers I, Tomlinson S R. The transcriptional foundation of pluripotency [J]. Development, 2009, 136(14): 2311-2322.
[16] Liu C, Lu Y, Wang B, et al. Clinical implications of stem cell gene Oct-4 expression in breast cancer [J]. Annals of surgery, 2011, 253(6): 1165-1171.
[17]Miyazawa K, Tanaka T, Nakai D, et al. Immunohistochemical expression of four different stem cell markers in prostate cancer: High expression of NANOG in conjunction with hypoxia?inducible factor?1α expression is involved in prostate epithelial malignancy [J]. Oncology letters, 2014, 8(3): 985-992.
[18]Chen Y C, Hsu H S, Chen Y W, et al. Oct-4 expression maintained cancer stem-like properties in lung cancer-derived CD133-positive cells [J]. PloS one, 2008, 3(7): e2637.
[19]Huang Z J, You J, Luo W Y, et al. Reduced tumorigenicity and drug resistance through the downregulation of octamer-binding protein 4 and Nanog transcriptional factor expression in human breast stem cells [J]. Molecular medicine reports, 2015, 11(3): 1647-1654.
[20]Koo B S, Lee S H, Kim J M, et al. Oct4 is a critical regulator of stemness in head and neck squamous carcinoma cells [J]. Oncogene, 2015, 34(18): 2317.
[21]Boumahdi S, Driessens G, Lapouge G, et al. SOX2 controls tumour initiation and cancer stem-cell functions in squamous-cell carcinoma [J]. Nature, 2014, 511(7508): 246.









