Recombinant Mouse Mothers against decapentaplegic homolog 3 (Smad3)
-
中文名稱:小鼠Smad3重組蛋白
-
貨號:CSB-YP804860MO
-
規格:
-
來源:Yeast
-
其他:
-
中文名稱:小鼠Smad3重組蛋白
-
貨號:CSB-EP804860MO
-
規格:
-
來源:E.coli
-
其他:
-
中文名稱:小鼠Smad3重組蛋白
-
貨號:CSB-EP804860MO-B
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:小鼠Smad3重組蛋白
-
貨號:CSB-BP804860MO
-
規格:
-
來源:Baculovirus
-
其他:
-
中文名稱:小鼠Smad3重組蛋白
-
貨號:CSB-MP804860MO
-
規格:
-
來源:Mammalian cell
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Smad3; Madh3; Mothers against decapentaplegic homolog 3; MAD homolog 3; Mad3; Mothers against DPP homolog 3; mMad3; SMAD family member 3; SMAD 3; Smad3
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length of Mature Protein
-
表達區域:2-425
-
氨基酸序列SSILPFTPP IVKRLLGWKK GEQNGQEEKW CEKAVKSLVK KLKKTGQLDE LEKAITTQNV NTKCITIPRS LDGRLQVSHR KGLPHVIYCR LWRWPDLHSH HELRAMELCE FAFNMKKDEV CVNPYHYQRV ETPVLPPVLV PRHTEIPAEF PPLDDYSHSI PENTNFPAGI EPQSNIPETP PPGYLSEDGE TSDHQMNHSM DAGSPNLSPN PMSPAHNNLD LQPVTYCEPA FWCSISYYEL NQRVGETFHA SQPSMTVDGF TDPSNSERFC LGLLSNVNRN AAVELTRRHI GRGVRLYYIG GEVFAECLSD SAIFVQSPNC NQRYGWHPAT VCKIPPGCNL KIFNNQEFAA LLAQSVNQGF EAVYQLTRMC TIRMSFVKGW GAEYRRQTVT STPCWIELHL NGPLQWLDKV LTQMGSPSIR CSSVS
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Receptor-regulated SMAD (R-SMAD) that is an intracellular signal transducer and transcriptional modulator activated by TGF-beta (transforming growth factor) and activin type 1 receptor kinases. Binds the TRE element in the promoter region of many genes that are regulated by TGF-beta and, on formation of the SMAD3/SMAD4 complex, activates transcription. Also can form a SMAD3/SMAD4/JUN/FOS complex at the AP-1/SMAD site to regulate TGF-beta-mediated transcription. Has an inhibitory effect on wound healing probably by modulating both growth and migration of primary keratinocytes and by altering the TGF-mediated chemotaxis of monocytes. This effect on wound healing appears to be hormone-sensitive. Regulator of chondrogenesis and osteogenesis and inhibits early healing of bone fractures. Positively regulates PDPK1 kinase activity by stimulating its dissociation from the 14-3-3 protein YWHAQ which acts as a negative regulator.
-
基因功能參考文獻:
- Data indicate E4BP4/NFIL3 as a direct Smad3 target gene critical for NK cell differentiation. PMID: 28262747
- Data suggest that Smad-specific E3 ubiquitin ligase 2 (SMURF2)-mediated SMAD3 protein (SMAD3) monoubiquitination interferes with the formation of a SMAD3-vitamin D receptor (VDR) complex. PMID: 28216630
- These data indicate that SMAD3- and beta-catenin-dependent induction occurs in the taurine transporter knockout mouse PMID: 28849477
- Cav-3 and Smad3 may be involved in the occurrence and development of viral myocarditis. PMID: 28770956
- In pancreata of mice and rats, TGFB promotes peripheral nociceptive sensitization via a direct effect on primary sensory neurons mediated by intra-neuronal SMAD3. PMID: 29505748
- We provide the evidence that over-expression of miR-145 could inhibit osteoclast differentiation, at least partially, by decreasing Smad3 expression PMID: 29577879
- conclude that VDD promotes tumor growth in the context of Smad3 disruption, potentially through regulation of TLR7 expression and beta-catenin activation PMID: 27456065
- SMAD3 is regulated by miR-489 in pulmonary fibrosis.miR-489 suppresses fibroblast differentiation by targeting Smad3. PMID: 27506999
- CRP is pathogenic in type-2 diabetes (T2DN). CRP may promote CD32b- NF-kappaB signaling to mediate renal inflammation; whereas, CRP may enhance renal fibrosis in T2DN via CD32b-Smad3-mTOR signaling. PMID: 27221338
- A Smad3-PTEN regulatory loop controls proliferation and apoptotic responses to TGF-beta in mouse endometrium. PMID: 28524854
- These findings indicate that both systemic factors and intrinsic properties of skin cells contribute to enhanced wound healing and less inflammatory reaction observed in Smad3 knock-out mice. PMID: 28130754
- Data show that Smad3 knockdown attenuated the effect of activin A on IL-6 release PMID: 28515224
- Sirt1 reduced endoplasmic reticulum stress and apoptosis of brown adipocytes in vivo/in vitro by inhibiting Smad3/ATF4 signaling pathway. PMID: 28030827
- these findings support the notion that Smad3 has important tumor suppressor function for breast cancer PMID: 27588495
- Treatment with metformin suppressed CCl4-induced expression of transforming growth factor beta 1 (TGF-beta1) and phosphorylation of Smad3. PMID: 28390311
- HSF1 activity is decreased in fibrotic hearts. HSF1 inhibits phosphorylation and nuclear distribution of Smad3 via direct binding to Smad3. Active Smad3 blocks the anti-fibrotic effect of HSF1. PMID: 28091697
- The results provide the first evidence that upregulation of TGFb/Smad3 in injured arteries induces local smooth muscle cells CXCR4 expression and cell migration, and consequently intimal hyperplasia. PMID: 27340942
- M1 macrophages with inhibited STAT3, STAT6 and/or SMAD3 effectively restrict tumor growth. The findings justify the development of new anti-tumor cell therapy technology. PMID: 29215829
- Aberrant Smad3 phosphoisoforms in cyst-lining epithelial cells in the cpk mouse suggest a qualitative rather than a quantitative abnormality of the TGF-beta/Smad3 pathway is involved in autosomal recessive polycystic kidney disease. PMID: 28877884
- The Smad3 and Bmal1 regulate p21 and S100A4 expression in myocardial stromal fibroblasts through TNF-alpha. PMID: 28721450
- conclude that the cardiovascular manifestations of Smad3 deficient mice are strain-specific, with myocyte necrosis in 129 mice and aortic rupture in C57BL/6J mice PMID: 29073282
- Smad3 binding to the -335 hypoxia-responsive element of the COL1A2 promoter required HIF-1alpha both in vitro and in kidney lysate from the disease model, suggesting formation of an HIF-1alpha-Smad3 transcriptional complex. Thus, HIF-1alpha-Smad3 has a novel interaction in glomerulosclerosis. PMID: 27503806
- Unexpectedly, a complex damage signal promotes co-localization of NF-kappaB, Smad3, and Nrf2 at Rev-erb-sensitive enhancers and drives expression of genes characteristic of multiple polarization states in the same cells. PMID: 27462873
- Store-operated calcium entry via Orai1 in mesangial cells negatively regulates the TGF-beta1/Smad3 signaling pathway. PMID: 28637791
- The results uncover an important aspect of the cross-talk between TGFbeta and Hippo signaling, showing that TGFbeta induces TAZ via a Smad3-independent, p38- and MRTF-mediated and yet MRTF translocation-independent mechanism. PMID: 28739802
- Data suggest that Gas5 suppresses Tgfb1/Smad3 signaling in vascular smooth muscle cell differentiation from mesenchymal progenitor cells; Gas5 competitively binds Smad3 via multiple RNA Smad-binding elements. (Gas5 = growth arrest-specific 5 long non-coding RNA; Tgfb1 = transforming growth factor beta 1; Smad3 = MAD homolog 3 protein) PMID: 28659340
- point mutant that was unable to bind pSMAD3 proved ineffective. These findings indicate that specifically targeting pSMAD3 can ameliorate both the direct and indirect fibroproliferative actions of TGF-beta. PMID: 28530637
- Soluble epoxide hydrolase inhibitor AUDA decreases bleomycin-induced pulmonary toxicity in mice by inhibiting the p38/Smad3 signaling pathway. PMID: 28694203
- A critical role for the Smad3-c-Jun pathway in the regulation of Fstl1. PMID: 28495857
- Mirtazapine suppressed 5-HT-mediated TGF-beta1/Smad3 and ERK1/2 signaling pathways as well as oxidative stress that contribute to the progression of liver fibrosis. PMID: 28623179
- These results suggested that TGF-beta1/Smad3 signaling was activated during CCl4-induced acute liver injury in mice, and Smad3 overexpression aggravated acute liver injury by promoting inflammatory cells infiltration, inflammatory cytokines release and hepatocytes apoptosis. PMID: 27224286
- gonadotropin-releasing hormone receptor mRNA levels were significantly elevated in knock-outs in both sexes. Interestingly, luteinizing hormone production was altered in a sex-specific fashion. Overall, our analyses demonstrate that SMAD3 is required for FSH synthesis in vivo PMID: 27994055
- Evaluated the effects of the loss of Smad3 on the development of experimental argon laser-induced choroidal neovascularization (CNV). The size of the CNV induced was significantly smaller in KO mice as compared with WT mice at day 14. PMID: 26950486
- emodin was found to increase the expression level of Sirt1, which decreased the level of deacetylated Smad3 to attenuate collagen deposition. Furthermore, the data suggested that there was direct binding between emodin and Sirt1. Sirt1regulated TGFb1/Smad signaling was involved in silica inhalationinduced lung fibrosis. PMID: 27748907
- HSP27 expression is upregulated in lung fibroblasts during pulmonary fibrosis, and subsequently, HSP27 modulates lung fibroblast differentiation through the Smad3 and ERK pathways. PMID: 27909724
- miR221 targets HMGA2 to inhibit leomycininduced pulmonary fibrosis through the TGFbeta1/Smad3 signaling pathway. PMID: 27513632
- Notch3 is an important protective factor for cardiac fibrosis in a myocardial infarction model, and the protective effect of Notch3 is attributable to its action on TGF-beta1/Smad3 signaling. PMID: 26487518
- Smad3 deficiency leads to aortic aneurysms and sudden death in the Smad3 knockout animal model. PMID: 27688095
- TGF-beta1-induced inhibition of PPARgamma transcription depends on formation of a functional transcriptional regulatory complex that includes Smad3, mSin3A and HDAC1 at the PPARgamma promoter. PMID: 27941310
- Activation of CB2 ameliorates myocardial fibrosis via Nrf2-mediated inhibition of TGF-beta1/Smad3 pathway in mice with myocardial infarction. PMID: 27614871
- The findings indicate that active Yap1 promotes the self-renewal of breast Tumor initiating cells by inhibiting Smad3 signaling. PMID: 26695440
- This study showed that Smad3 deficiency inhibits LTP induction by enhancing phasic and tonic GABAA receptor-mediated neurotransmission. PMID: 26826552
- Suggest that hyperoxia-increased high-tidal-volume ventilation-induced acute lung injury partially depends on the Src and Smad3 pathways. PMID: 26377087
- Smad3 is a key transcriptional factor of TGF-beta signaling that differentially regulates T cell immune responses in a mouse model of cardiac allograft rejection PMID: 26219259
- Smad3 deficiency leads to mandibular condyle degradation via the sphingosine 1-phosphate (S1P)/S1P3 signaling axis PMID: 26272361
- pSmad3L-Ser signalling correlates with carcinogenesis of colon tumors and this work supports the hypothesis that pSmad2/3L-Thr immunostaining-positive cells are cancer PMID: 25908723
- SMAD3 deficiency promotes vessel wall remodeling, collagen fiber reorganization and leukocyte infiltration in an inflammatory abdominal aortic aneurysm mouse model. PMID: 25985281
- Gremlin or Ad5.TGFbeta2 elevate IOP and upregulate the ECM protein FN in the TM of mice. These data show that gremlin signals through the Smad3-dependent pathway in the TM to elevate IOP. PMID: 26284554
- PTEN loss initiates tubular dysfunction via SMAD3- and p53-mediated fibrotic gene induction, with accompanying PAI-1-dependent proliferative arrest, and cooperates with TGFbeta1 to induce the expression of profibrotic genes and tubular apoptosis. PMID: 25810340
- Smad3 mediates diabetic cardiac hypertrophy, fibrosis, and diastolic dysfunction, while preserving normal cardiac geometry and maintaining the integrity of the vascular wall. PMID: 25985794
顯示更多
收起更多
-
亞細胞定位:Cytoplasm. Nucleus.
-
蛋白家族:Dwarfin/SMAD family
-
組織特異性:Highly expressed in the brain and ovary. Detected in the pyramidal cells of the hippocampus, granule cells of the dentate gyrus, granular cells of the cerebral cortex and the granulosa cells of the ovary.
-
數據庫鏈接:
Most popular with customers
-
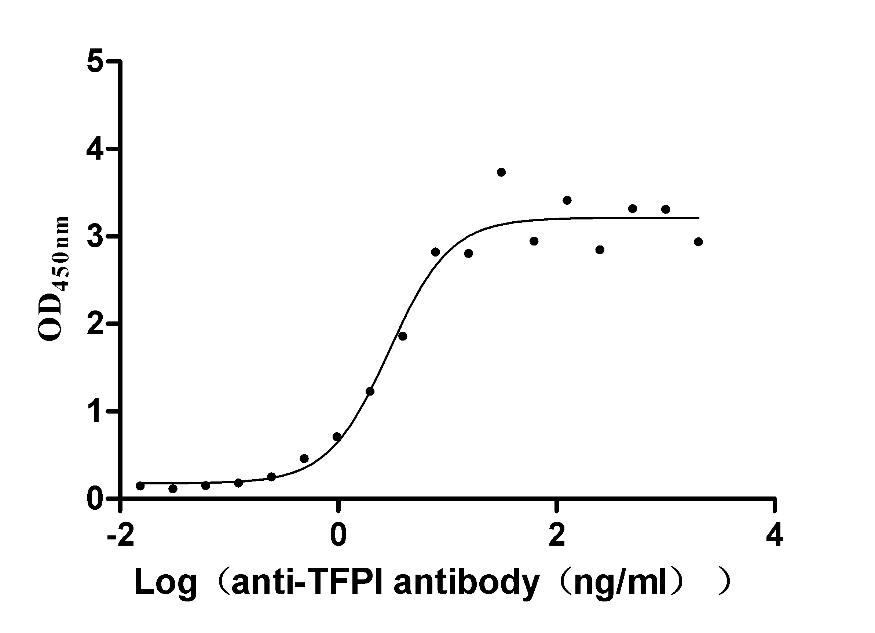
Recombinant Rabbit Tissue factor pathway inhibitor (TFPI) (Active)
Express system: Mammalian cell
Species: Oryctolagus cuniculus (Rabbit)
-
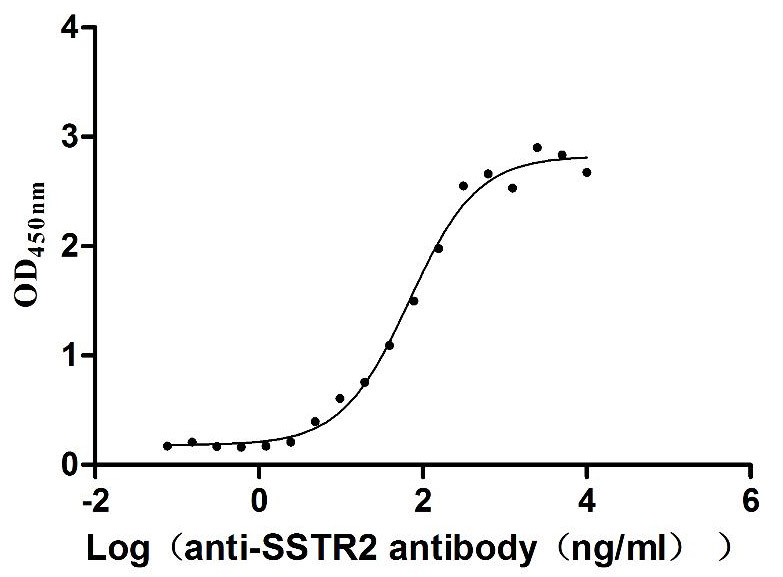
Recombinant Human Somatostatin receptor type 2 (SSTR2)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
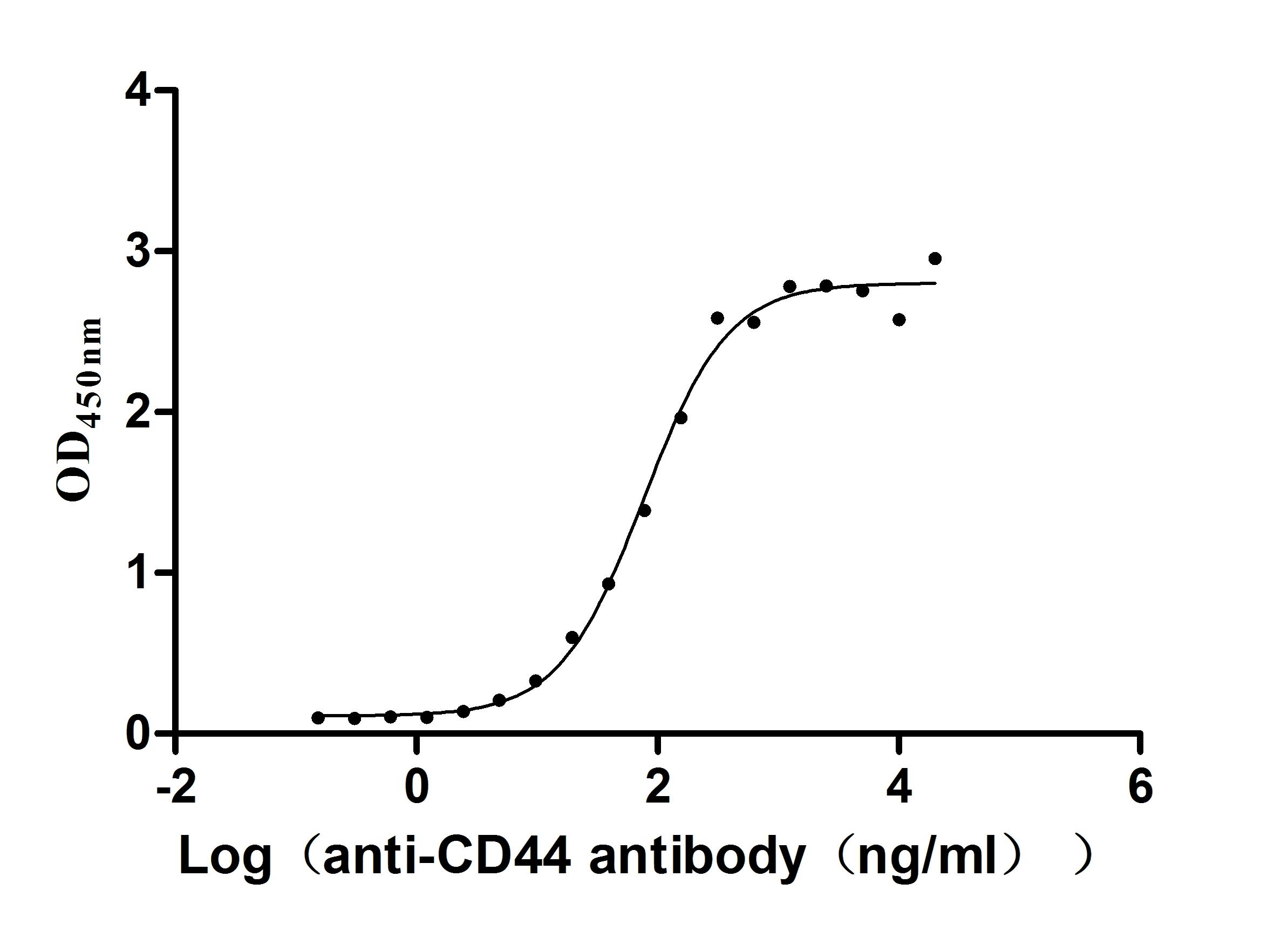
Recombinant Macaca fascicularis CD44 antigen (CD44), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
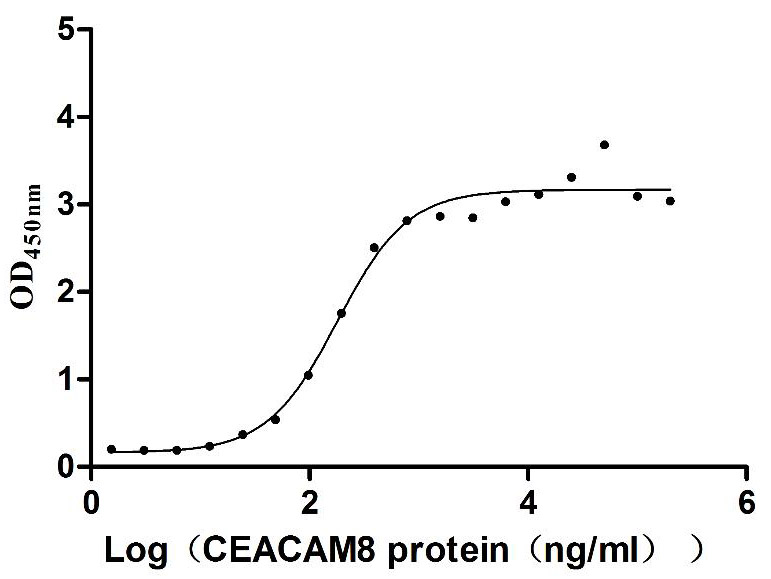
Recombinant Human Carcinoembryonic antigen-related cell adhesion molecule 6 (CEACAM6) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
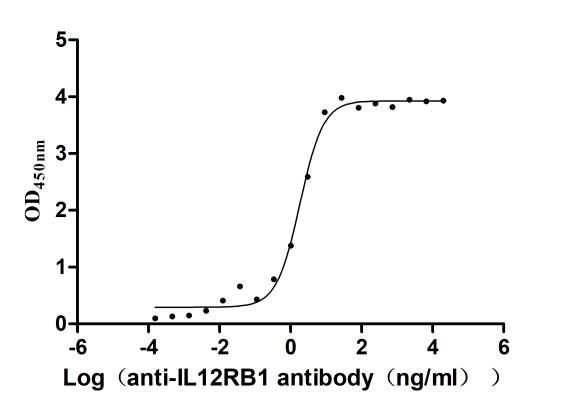
Recombinant Human Interleukin-12 receptor subunit beta-1(IL12RB1),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









