Recombinant Mouse Catenin alpha-1 (Ctnna1), partial
-
中文名稱:Recombinant Mouse Catenin alpha-1(Ctnna1) ,partial
-
貨號:CSB-YP006164MO
-
規格:
-
來源:Yeast
-
其他:
-
中文名稱:Recombinant Mouse Catenin alpha-1(Ctnna1) ,partial
-
貨號:CSB-EP006164MO
-
規格:
-
來源:E.coli
-
其他:
-
中文名稱:Recombinant Mouse Catenin alpha-1(Ctnna1) ,partial
-
貨號:CSB-EP006164MO-B
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Mouse Catenin alpha-1(Ctnna1) ,partial
-
貨號:CSB-BP006164MO
-
規格:
-
來源:Baculovirus
-
其他:
-
中文名稱:Recombinant Mouse Catenin alpha-1(Ctnna1) ,partial
-
貨號:CSB-MP006164MO
-
規格:
-
來源:Mammalian cell
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Ctnna1; Catna1; Catenin alpha-1; 102 kDa cadherin-associated protein; Alpha E-catenin; CAP102
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Partial
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Associates with the cytoplasmic domain of a variety of cadherins. The association of catenins to cadherins produces a complex which is linked to the actin filament network, and which seems to be of primary importance for cadherins cell-adhesion properties. Can associate with both E- and N-cadherins. Originally believed to be a stable component of E-cadherin/catenin adhesion complexes and to mediate the linkage of cadherins to the actin cytoskeleton at adherens junctions. In contrast, cortical actin was found to be much more dynamic than E-cadherin/catenin complexes and CTNNA1 was shown not to bind to F-actin when assembled in the complex suggesting a different linkage between actin and adherens junctions components. The homodimeric form may regulate actin filament assembly and inhibit actin branching by competing with the Arp2/3 complex for binding to actin filaments. Involved in the regulation of WWTR1/TAZ, YAP1 and TGFB1-dependent SMAD2 and SMAD3 nuclear accumulation. May play a crucial role in cell differentiation.
-
基因功能參考文獻:
- alphaE-catenin is essential for inhibiting nuclear YAP localization and cell proliferation. This function of alphaE-catenin is required for formation of the tooth signalling centre, the enamel knot (EK), which maintains dental mesenchymal condensation and epithelial invagination. alphaE-catenin restricts YAP/TAZ activity to establish a group of non-dividing and specialized cells that constitute a signalling centre. PMID: 27405641
- Catna1 functions as a positive regulator of pancreatic islet cell lineage differentiation by repressing the sonic hedgehog pathway. PMID: 28793255
- alpha-catenins regulate intercalated disc maturation and actomyosin contractility, which, in turn, control Yap subcellular localization, thus providing an explanation for the loss of proliferative capacity in the newborn mammalian heart. PMID: 29467248
- In this study, the authors revealed how alpha-catenin retains its activated state while avoiding unfolding under tension. PMID: 27109499
- cadherin 2 (CDH2) and CDH4 cooperate to regulate radial migration in mouse brain via the protein tyrosine phosphatase 1B (PTP1B) and alpha- and beta-catenins. PMID: 27151949
- alphaT-catenin is a constitutively active actin-binding protein that can physically couple the cadherin.catenin complex to F-actin in the absence of tension. PMID: 27231342
- Two Dtna interactors, alpha-catulin (phosphorylation independent) and Grb2 (phosphorylation dependent) are localized to neuromuscular junctions in vivo, and are required for proper organization of neurotransmitter receptors on myotubes. PMID: 26769899
- alphaE-catenin inhibits beta4 integrin-mediated activation of SRC tyrosine kinase PMID: 27013234
- E-cadherin/alphaE-catenin chimeras used previously do not mimic alphaE-catenin in the native CCC, and imply that both CCC-bound monomer and cytosolic homodimer alphaE-catenin are required for strong cell-cell adhesion. PMID: 26416960
- Loss of alpha-catenin elicits a cholestatic response and impairs liver regeneration PMID: 25355493
- alphaE-catenin binding to filamentous actin favors assembly of unbranched filament bundles that are protected from severing over more dynamic, branched filament arrays. PMID: 24068324
- Fusing VE-cadherin to alpha-catenin presents a mutation, which leads to embryonic lethality, due to a lack of fetal liver hematopoiesis and severe lymphedema but no detectable defects in blood vessel formation and remodeling. PMID: 24567373
- alphaE-catenin recruits vinculin to adherens junctions more effectively than alphaN-catenin, partly because of its higher affinity for actin filaments. PMID: 23589308
- our results show that alpha-catenin-mediated cell adhesion and cell organization are important for the fissure closure in mice, and further suggest that genes that regulate cell adhesion may underlie certain coloboma cases in humans PMID: 23240058
- alpha-Catenin and vinculin cooperate to promote high E-cadherin-based adhesion strength PMID: 23266828
- alpha-catenin, previously implicated in tumor suppression and cell density sensing in the skin, is an upstream negative regulator of Yap1; alpha-catenin controls Yap1 activity and phosphorylation by modulating its interaction with 14-3-3 and PP2A phosphatase. PMID: 21376238
- Data show that alphaE-catenin D3 binds strongly to vinculin, whereas larger fragments and full-length alphaE-catenin bind approximately 1,000-fold more weakly. PMID: 22586082
- An E-cadherin-alpha-catenin fusion protein (Ealpha) restored full cell-adhesion function and organized the actin-based cytoskeleton and ZO-1, an actin filament binding protein, in F9 cells lacking all endogenous cadherin-catenin complex components. PMID: 21685705
- An evolutionarily conserved PTEN-C/EBPalpha-CTNNA1 axis controls myeloid development and transformation. PMID: 20371743
- In the cortex, alpha-catenin is switched from alphaE-catenin in the VZ to alphaN-catenin II in the intermediate zone (IMZ). PMID: 16185771
- These results suggest that the linkage between the cadherin-catenin complex and actin filaments is more dynamic than previously appreciated. PMID: 16325582
- Data show that alpha-catenin exists as a monomer or a homodimer, with the monomer binding more strongly to E-cadherin-beta-catenin and the dimer preferentially binding actin filaments. PMID: 16325583
- Reductions in alpha-catenin, activation of NF-kappaB, and inflammation are common features of human squamous cell carcinomas of the PMID: 16452166
- alphaE-catenin is highly expressed in neuroepithelial precursor cells in the developing cortical ventricular zone, with markedly reduced expression in the cortical plate PMID: 16457793
- The cadherin-11 cytoplasmic juxtamembrane domain is a regulator of alpha-catenin turnover at adherens junctions and actin-cytoskeletal organization that is critical for intercellular motility and rearrangement in multicellular clusters. PMID: 16525026
- proposed that alphaE-catenin connects cell-density-dependent adherens junctions with the developmental hedgehog pathway and that this connection may provide a negative feedback loop controlling the size of developing cerebral cortex PMID: 16543460
- Ablation of alpha-E-catenin leads to defects in cardiomyocyte structural integrity that result in unique forms of cardiomyopathy and predisposed susceptibility to death after myocardial stress. PMID: 16923756
- Endogenous alphaE-catenin has no significant impact on beta-catenin transcriptional activities in the developing mammalian brain. PMID: 18397997
- Alpha-E-catenin functions as a dynamic link between the dynactin complex and actin and, thus, integrates the microtubule and actin cytoskeleton during intracellular trafficking. PMID: 19075109
- These results suggest that alpha-catenin overexpression increases osteoblastic differentiation by increasing cell-cell adhesion rather than Wnt-/beta-catenin-signaling. PMID: 19324011
- Focal reduction of Ctnna1 causes premature differentiation and reduction of beta catenin signaling during cortical development. PMID: 19389371
顯示更多
收起更多
-
亞細胞定位:Cytoplasm, cytoskeleton. Cell junction, adherens junction. Cell membrane; Peripheral membrane protein; Cytoplasmic side. Cell junction. Note=Found at cell-cell boundaries and probably at cell-matrix boundaries.
-
蛋白家族:Vinculin/alpha-catenin family
-
組織特異性:Expressed ubiquitously in normal tissues.
-
數據庫鏈接:
Most popular with customers
-
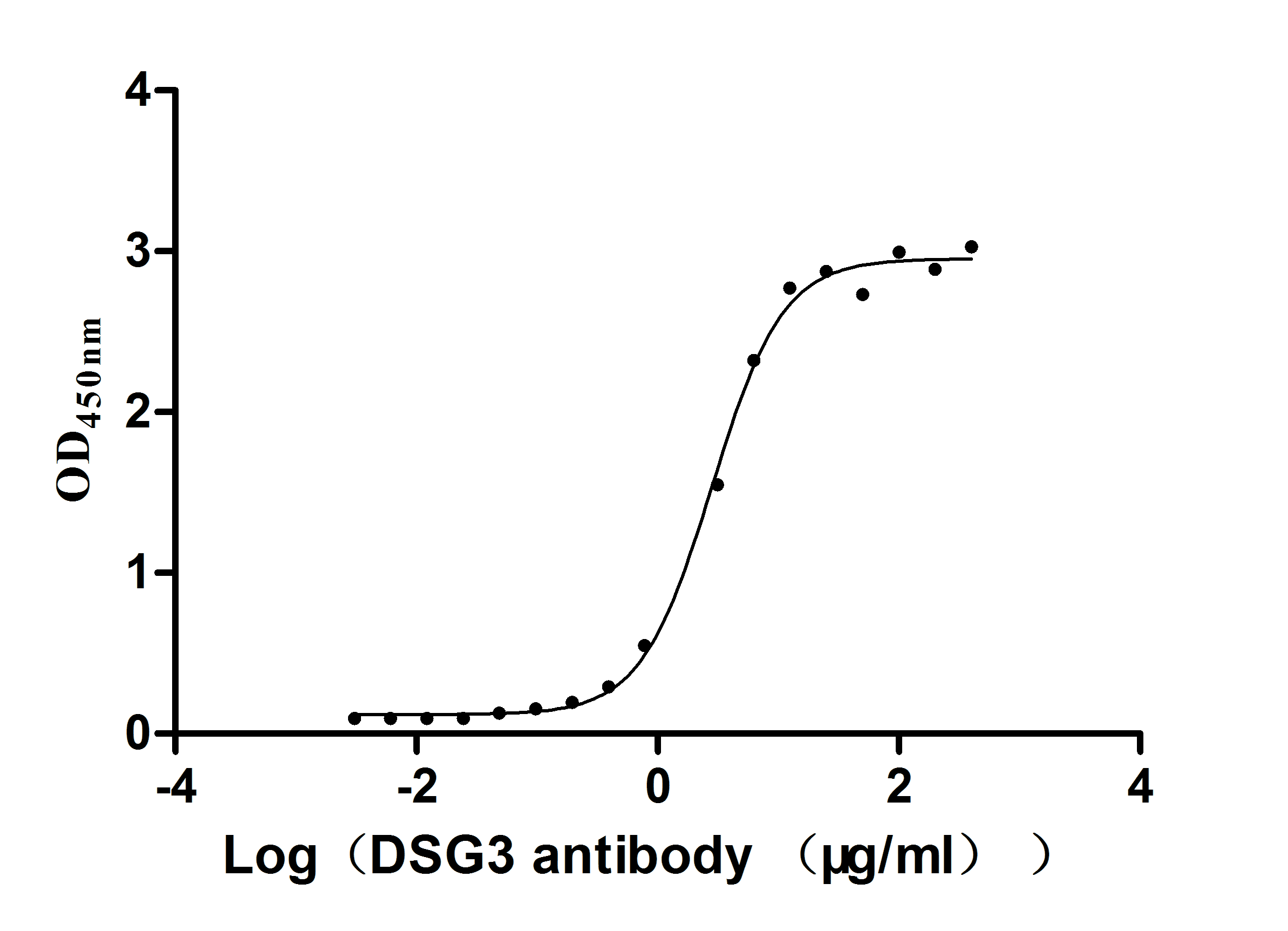
Recombinant Mouse Desmoglein-3 (Dsg3), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
-AC1.jpg)
Recombinant Human Poliovirus receptor (PVR) (I340M), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
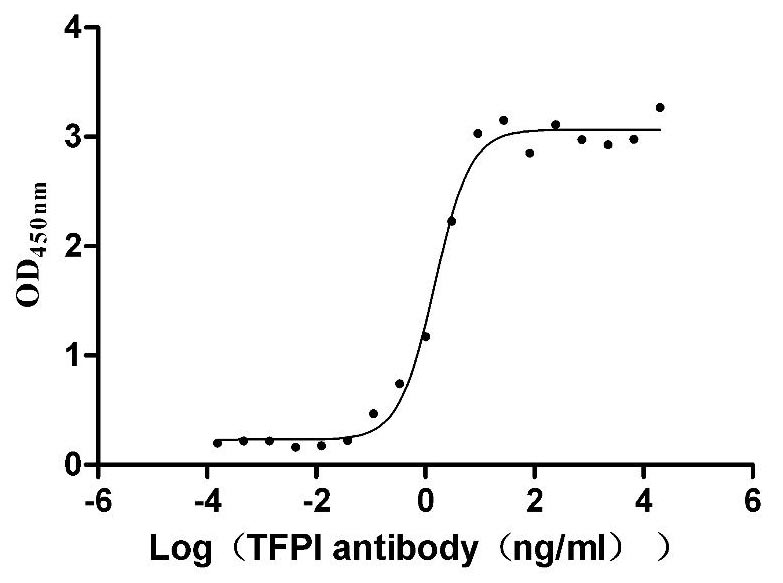
Recombinant Human Tissue factor pathway inhibitor (TFPI), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
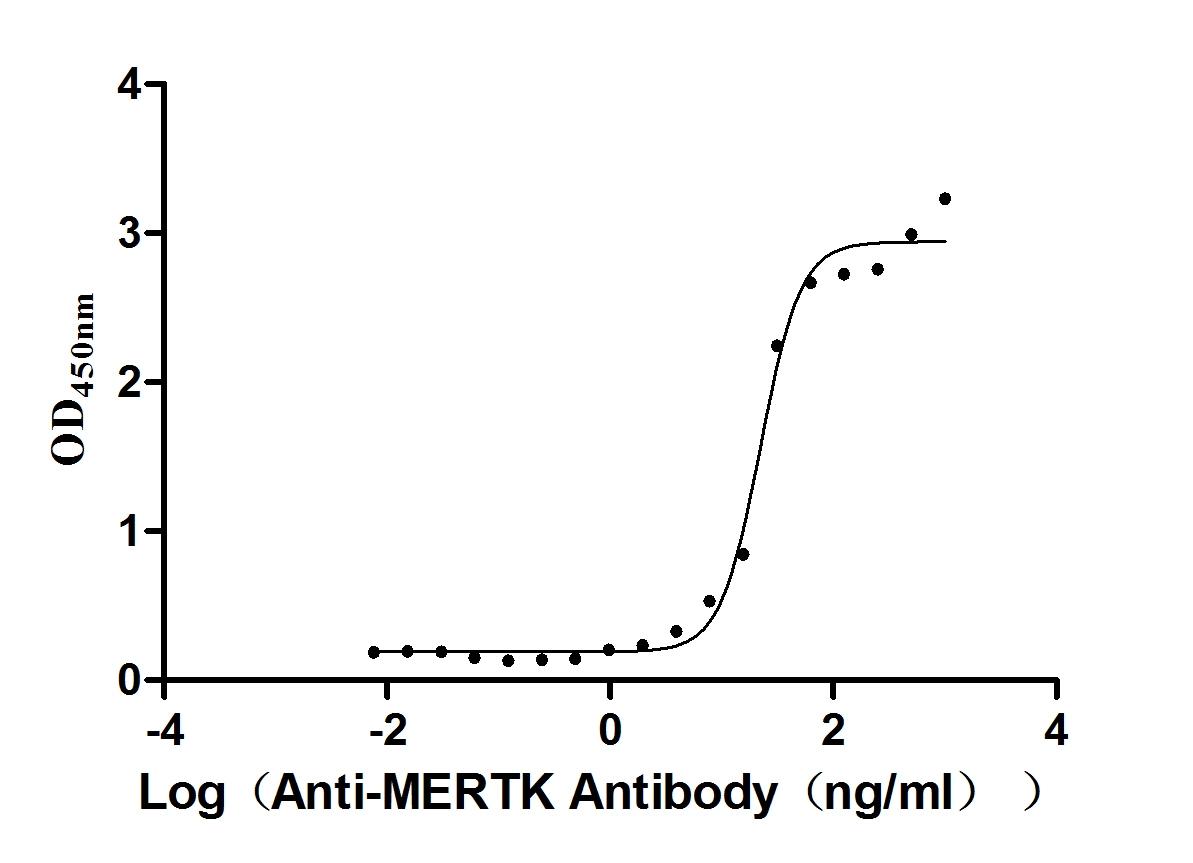
Recombinant Mouse Tyrosine-protein kinase Mer (Mertk), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
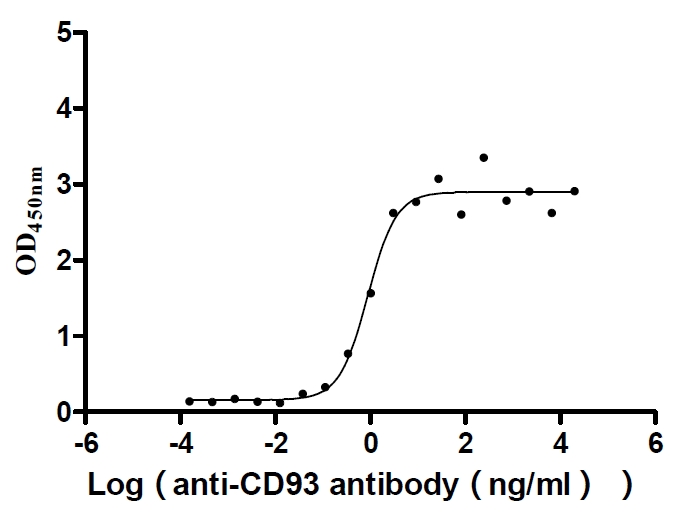
Recombinant Human Complement component C1q receptor (CD93), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human IL12B&IL12A Heterodimer Protein (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Transferrin receptor protein 1 (TFRC), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Myosin regulatory light chain 12B (MYL12B) (Active)
Express system: E.coli
Species: Homo sapiens (Human)









