Recombinant Human Interferon regulatory factor 8 (IRF8 ICSBP1)
-
中文名稱:
-
貨號:CSB-YP011823HU
-
規格:
-
來源:Yeast
-
其他:
-
中文名稱:
-
貨號:CSB-EP011823HU
-
規格:
-
來源:E.coli
-
其他:
-
中文名稱:
-
貨號:CSB-EP011823HU-B
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:
-
貨號:CSB-BP011823HU
-
規格:
-
來源:Baculovirus
-
其他:
-
中文名稱:
-
貨號:CSB-MP011823HU
-
規格:
-
來源:Mammalian cell
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Interferon regulatory factor 8; IRF-8; Interferon consensus sequence-binding protein; H-ICSBP; ICSBP; IRF8 ICSBP1
-
種屬:Homo sapiens (Human)
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Transcription factor that specifically binds to the upstream regulatory region of type I interferon (IFN) and IFN-inducible MHC class I genes (the interferon consensus sequence (ICS)). Can both act as a transcriptional activator or repressor. Plays a negative regulatory role in cells of the immune system. Involved in CD8(+) dendritic cell differentiation by forming a complex with the BATF-JUNB heterodimer in immune cells, leading to recognition of AICE sequence (5'-TGAnTCA/GAAA-3'), an immune-specific regulatory element, followed by cooperative binding of BATF and IRF8 and activation of genes. Required for the development of plasmacytoid dendritic cells (pDCs), which produce most of the type I IFN in response to viral infection. Positively regulates macroautophagy in dendritic cells.
-
基因功能參考文獻:
- Exogenous expression of IRF8 in the silenced or downregulated lung cancer cell lines restored the sensitivity of lung cancer cells to apoptosis, and arrested cells at the G0/G1 phase. IRF8 bound to the T-cell factor/lymphoid enhancer factor (TCF /LEF) promoter, thus repressing beta-catenin nuclear translocation and its activation. PMID: 30466106
- Study shows that hypermethylation of IRF8 associated with decreased mRNA expression in dendritic cells confers risk to VKH disease. PMID: 28432342
- IRF8 upregulation in tumor cells inhibited the generation of Th17 cells in vitro, and this may be mediated by the downregulation of RORgammat. we found that a high level of IRF8 in the DLBCL tumor microenvironment was a predictor of poor survival in DLBCL patients. PMID: 28537908
- These findings identify IRF8 as a novel tumor suppressor regulating IFN-gamma/STAT1 signaling and beta-catenin signaling in breast cancer. PMID: 28388578
- Irf8 induction, but not its knockdown, decreased APL leukemogenic potential through driving monocytic maturation. PMID: 28492552
- Total cellular protein presence of the transcription factor IRF8 does not necessarily correlate with its nuclear presence. PMID: 27582125
- IRF8 is dispensable for induced pluripotent stem cell and embryonic stem cell differentiation into hemogenic endothelium and for endothelial-to-hematopoietic transition. PMID: 28090699
- TP(thymidine phosphorylase ) curbed the expression of three proteins-IRF8, RUNX2, and osterix. This downregulation was epigenetically driven: High levels of 2DDR, a product of TP secreted by myeloma cells, activated PI3K/AKT signaling and increased the methyltransferase DNMT3A's expression PMID: 27658717
- It findings provide evidence for an additional mechanism of epigenetic IRF8 silencing during osteoclastogenesis that likely works cooperatively with DNA methylation, further emphasizing the importance of IRF8 as a negative regulator of osteoclastogenesis. PMID: 27183582
- Data suggest that ubiquitin specific protease 4 (USP4) interacts with interferon regulatory factor 8 (IRF8) and, by its Lys48-specific deubiquitinase/endopeptidase activity, stabilizes IRF8 protein levels in regulatory T-lymphocytes; USP4 and IRF8 are also expressed in helper T-lymphocytes. PMID: 28477415
- these data indicate that human NK cells require IRF8 for development and functional maturation and that dysregulation of this function results in severe human disease, thereby emphasizing a critical role for NK cells in human antiviral defense. PMID: 27893462
- down-regulation of IRF8 in the wound leads to impaired wound healing possibly through the regulation of macrophage function and apoptosis in skin wound. PMID: 27709315
- IRF8 may contribute to the genetic susceptibility of Behcet's disease by regulating IRF8 expression and cytokine production. PMID: 26794091
- IRF5 and IRF8, two transcription factors with opposing functions, control TLR9 signaling in human plasmacytoid dendritic cells. PMID: 26613957
- cytarabine-induced upregulation of the IRF8 in leukemic cells involves increased levels of ZNF224, which can counteract the repressive activity of WT1 on the IRF8-promoter PMID: 26563595
- Expression of WT1and IRF8 showed a moderate inverse correlation in acute myeloid leukemia patients. WT1 can be used as an minimal residual disease marker, especially in patients without recurrent genetic abnormalities. PMID: 25387409
- Results show that IRF8 is a possible genetic variant associated with the development of HT and production of thyroid antibody PMID: 25989711
- The IRF8 gene variant influenced the interaction between IRF8 and NF-kappaB and thus susceptibility to systemic sclerosis. PMID: 25880423
- This article provides an overview of recent advances in our understanding of the role of IRF8 in myelopoiesis and related diseases. [review] PMID: 25749660
- This study demonstrated that the Polymorphism, Single Nucleotide of IRF8 is associated with multiple sclerosis in woman in Russia. PMID: 25903733
- gene variants in IRF5, IRF8 and GPC5 were not associated with risk of relapse or disease progression in multiple sclerosis PMID: 24943672
- Irf8 forms a negative feedback loop with Cebpb, a monocyte-derived DC epigenetic fate-determining transcription factor. PMID: 25453760
- MN1 prevents activation of the immune response pathway, and suggest restoration of IRF8 signaling as therapeutic target in AML PMID: 24957708
- Data demonstrate that IRF8 as a functional tumor suppressor is frequently methylated in RCC, and IRF8-mediated interferon signaling is involved in RCC pathogenesis. PMID: 25109451
- The IRF8 gene polymorphisms might be associated with susceptibility to SLE. PMID: 24034601
- Data demonstrate a novel function for ICSBP in epithelial-mesenchymal transition, cell motility, and invasion through the TGF-beta and Snail signaling pathways. PMID: 24832596
- These data reveal a previously unrecognized BCR-ABL-STAT5-IRF-8 network, which widens the repertoire of potentially new anti-chronic myeloid leukemia targets. PMID: 24753251
- IRF8 is associated with germinal center B-cell-like type of diffuse large B-cell lymphoma and exceptionally involved in translocation t(14;16)(q32.33;q24.1). PMID: 23573829
- Mutations in linker histone genes HIST1H1 B, C, D, and E; OCT2 (POU2F2); IRF8; and ARID1A underlying the pathogenesis of follicular lymphoma. PMID: 24435047
- the prognostic significance of IRF8 transcripts in other populations of AML patients PMID: 23967110
- In a Korean population, there was no association of IRF8 polymorphisms, with inflammatory demyelinating diseases. PMID: 22994200
- novel insights into the contribution of EBNA3C to EBV-mediated B-cell transformation through regulation of IRF4 and IRF8 and add another molecular link to the mechanisms by which EBV dysregulates cellular activities PMID: 23658517
- Using a capture sequencing strategy, we discovered the B-cell relevant genes IRF8, EBF1, and TNFSF13 as novel targets for IGH deregulation. PMID: 23775715
- Coronary heart disease in systemic lupus erythematosus is associated with IRF8 gene variants. PMID: 23661672
- A 3'UTR variant in IRF8 gene is associated with risk of chronic lymphocytic leukemia. PMID: 23307532
- these results underscore a key role of IRF-8 in the cross talk between melanoma and immune cells, thus revealing its critical function within the tumor microenvironment in regulating melanoma progression and invasiveness. PMID: 23308054
- IRF8 is required for ontogeny of the myeloid lineage and for host response to mycobacteria. The K108E & T80A mustations cause distinct forms of a novel primary immunodeficiency & associated susceptibility to mycobacteria. Review. PMID: 23468103
- The association of IRF1 and IRF8 variants with tuberculosis susceptibility was investigated. PMID: 22879909
- In resting macrophages, some IRF8 is conjugated to small ubiquitin-like modifiers (SUMO) 2/3 through the lysine residue 310. Macrophage activation prompts a reduction in SUMOylated IRF8. PMID: 22942423
- Four additional susceptibility loci (IRF8, TMEM39A, IKZF3, and ZPBP2) for systemic lupus erythematosus were robustly established a multiethnic population (European, African American, Asian, Hispanic, Gullah, and Amerindian). PMID: 22464253
- interaction between Tel and Tel-PdgfRbeta decreases Tel/Icsbp/Hdac3 binding to the PTPN13 cis element, resulting in increased transcription. PMID: 22262849
- Data showed that IRF8 target genes contributes to multiple aspects of the biology of mature B cells including critical components of the molecular crosstalk among GC B cells, T follicular helper cells, and follicular dendritic cells. PMID: 22096565
- Association analysis identified five SLE susceptibility genes reaching genome-wide levels of significance : NCF2 ,IKZF1 ,IRF8 ,IFIH1 , and TYK2 PMID: 22046141
- IRF4 has activities similar to IRF8 in regulating myeloid cell development PMID: 22003407
- [review] Induction of transcriptional repressors such as IRF8 is one of the mechanisms that inhibits osteoclastogenesis. PMID: 22082370
- study identifies a novel role for ICSBP in regulating cell growth via TGF-beta receptor upregulation and subsequent activation of the TGF-beta receptor/TAK-1/p38 pathway PMID: 21625229
- The promoter was methylated in many MDS or AML patients. This may be the main mechanism of ICSBP inactivation in myeloid malignancies & may be functionally important for accumulation of chromosome aberrations during leukemic progression. PMID: 21475251
- association of single nucleotide polymorphisms to multiple sclerosis PMID: 21552549
- We detected two distinct disease-causing mutations affecting interferon regulatory factor 8 (IRF8). Both mutations impair IRF8 transcriptional activity. PMID: 21524210
- Findings determine the mechanism of IRF8 downregulation in CML cells. PMID: 21487040
顯示更多
收起更多
-
相關疾病:Immunodeficiency 32A (IMD32A); Immunodeficiency 32B (IMD32B)
-
亞細胞定位:Nucleus. Cytoplasm.
-
蛋白家族:IRF family
-
組織特異性:Predominantly expressed in lymphoid tissues.
-
數據庫鏈接:
Most popular with customers
-
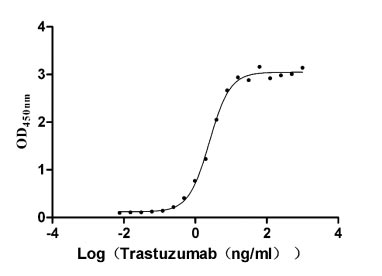
Recombinant Human Receptor tyrosine-protein kinase erbB-2 (ERBB2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
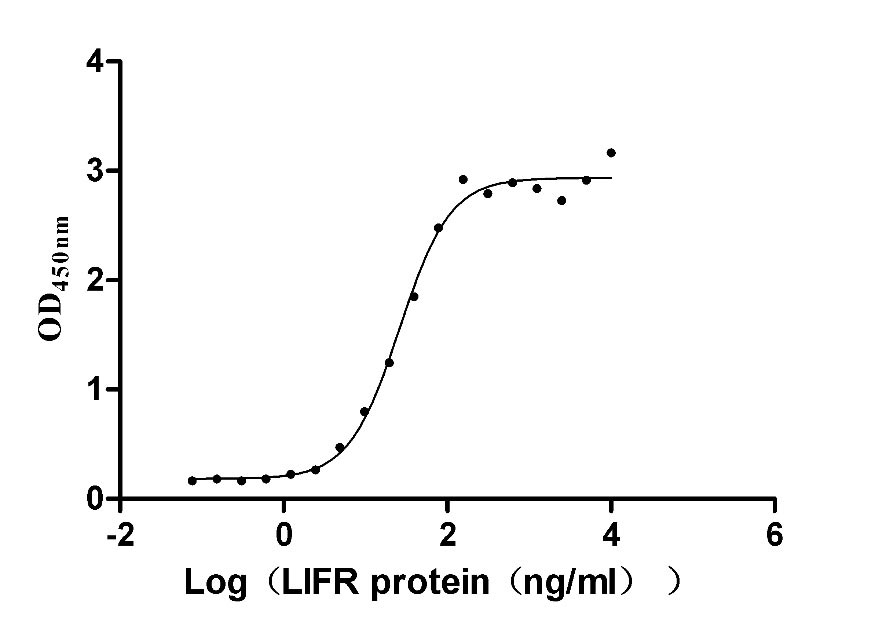
Recombinant Human Leukemia inhibitory factor (LIF) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Rat Intestinal-type alkaline phosphatase 1 (Alpi) (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
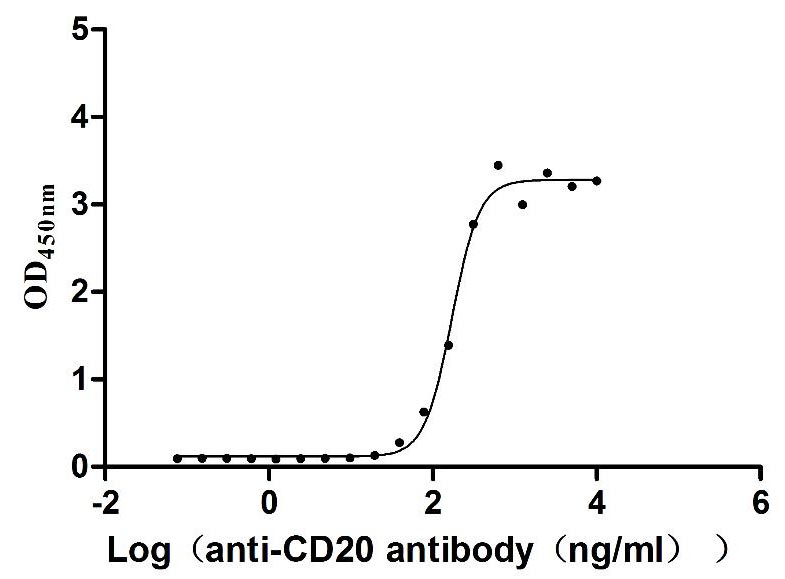
Recombinant Dog B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Canis lupus familiaris (Dog) (Canis familiaris)
-
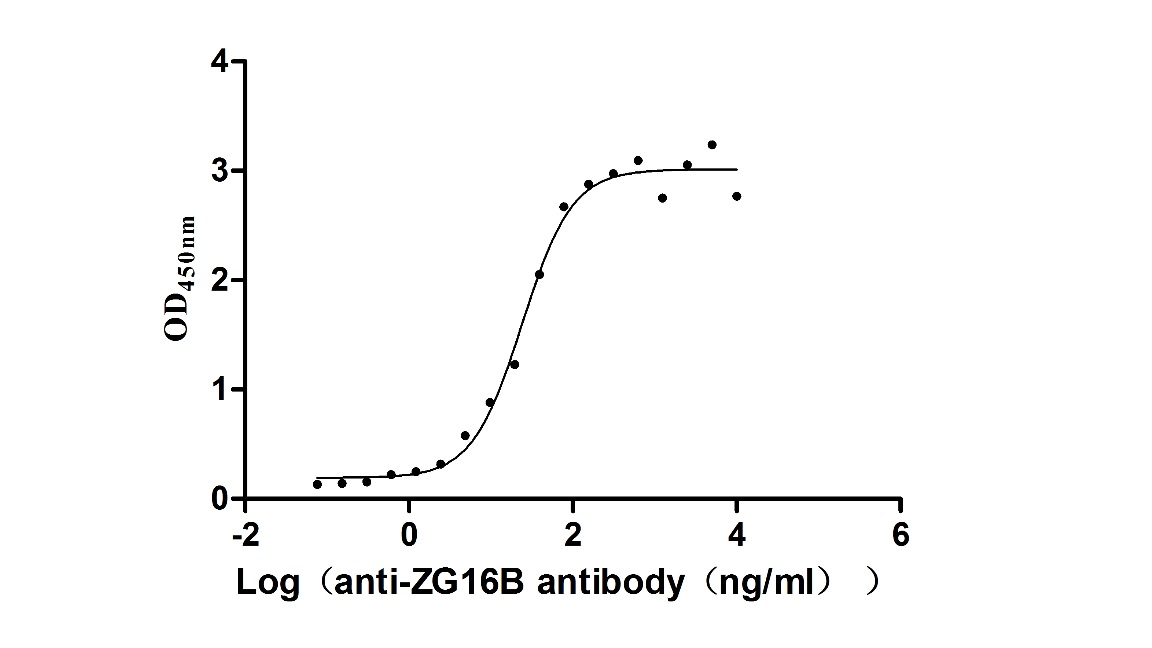
Recombinant Macaca fascicularis zymogen granule protein 16 homolog B (ZG16B) (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Human Cadherin-17 (CDH17), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Transmembrane 4 L6 family member 1(TM4SF1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Express system: Mammalian cell
Species: Homo sapiens (Human)









