Recombinant Human Histone deacetylase 5 (HDAC5), partial
-
中文名稱:人HDAC5重組蛋白
-
貨號:CSB-YP010241HU
-
規(guī)格:
-
來源:Yeast
-
其他:
-
中文名稱:人HDAC5重組蛋白
-
貨號:CSB-EP010241HU
-
規(guī)格:
-
來源:E.coli
-
其他:
-
中文名稱:人HDAC5重組蛋白
-
貨號:CSB-EP010241HU-B
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:人HDAC5重組蛋白
-
貨號:CSB-BP010241HU
-
規(guī)格:
-
來源:Baculovirus
-
其他:
-
中文名稱:人HDAC5重組蛋白
-
貨號:CSB-MP010241HU
-
規(guī)格:
-
來源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Antigen NY CO 9; Antigen NY-CO-9; HD5; HDAC 5; HDAC5; HDAC5_HUMAN; Histone deacetylase 5; NY CO 9
-
種屬:Homo sapiens (Human)
-
蛋白長度:Partial
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:Responsible for the deacetylation of lysine residues on the N-terminal part of the core histones (H2A, H2B, H3 and H4). Histone deacetylation gives a tag for epigenetic repression and plays an important role in transcriptional regulation, cell cycle progression and developmental events. Histone deacetylases act via the formation of large multiprotein complexes. Involved in muscle maturation by repressing transcription of myocyte enhancer MEF2C. During muscle differentiation, it shuttles into the cytoplasm, allowing the expression of myocyte enhancer factors. Involved in the MTA1-mediated epigenetic regulation of ESR1 expression in breast cancer. Serves as a corepressor of RARA and causes its deacetylation. In association with RARA, plays a role in the repression of microRNA-10a and thereby in the inflammatory response.
-
基因功能參考文獻(xiàn):
- These results demonstrate a previously unknown negative epigenetic regulation of hematopoietic stem cells (HSC) homing and engraftment by HDAC5, and allow for a new and simple translational strategy to enhance HSC transplantation. PMID: 30013077
- Collectively, these data indicate that vIRF3 alters global gene expression and induces a hypersprouting formation in an HDAC5-binding-dependent and lymphatic endothelial cell-specific manner, ultimately contributing to Kaposi's sarcoma-associated herpesvirus-associated pathogenesis. PMID: 29339432
- High HDAC5 expression is associated with invasion of lung cancer. PMID: 30066893
- HO-1 plays a key role in protecting tumor cells from apoptosis, in a process that involves Smad7 and HDAC4/5 in apoptosis of B-ALL cells PMID: 29886060
- These findings demonstrate a novel mechanism for deregulation of HDAC5 in non-small cell lung cancer (NSCLC)and suggest that miR5895p/HDAC5 pathway may represent a new prognostic biomarker and therapeutic target against NSCLC. PMID: 28440397
- HDAC5 was extensively expressed in human BC tissues, and high HDAC5 expression was associated with an inferior prognosis. PMID: 27177225
- HDAC5 is a negative predictor of disease-free and overall survival in pancreatic neuroendocrine tumor patients. PMID: 28235630
- Interference with both glucose and glutamine supply in HDAC5-inhibited cancer cells significantly increases apoptotic cell death. PMID: 28414307
- these results suggest that HDAC5 is critical in regulating LSD1 protein stability through post-translational modification, and the HDAC5-LSD1 axis has an important role in promoting breast cancer development and progression. PMID: 27212032
- The expression of HDAC5 was significantly increased in endothelial cells (ECs) from patients with systemic sclerosis (SSc) compared to healthy control endothelial cells. Silencing of HDAC5 in SSc ECs restored normal angiogenesis. HDAC5 knockdown followed by ATAC-seq assay in SSc ECs identified key HDAC5-regulated genes involved in angiogenesis and fibrosis, such as CYR61, PVRL2, and FSTL1. PMID: 27482699
- the migration and invasion of hepatocellular carcinoma cells were impaired by knockdown of histone deacetylase 5 or hypoxia-inducible factor-1alpha but rescued when eliminating homeodomain-interacting protein kinase-2 in hepatocellular carcinoma cells, which suggested the critical role of histone deacetylase 5-homeodomain-interacting protein kinase-2-hypoxia-inducible factor-1alpha pathway in hypoxia-induced metastasis. PMID: 28653891
- HDAC5 inhibits hepatic lipogenic genes expression by attenuating the transcriptional activity of liver X receptor. PMID: 27614433
- HDAC5 promotes cellular proliferation through the upregulation of cMet, and may provide a novel therapeutic target for the treatment of patients with Wilms' tumor. PMID: 26847592
- Formononetin-combined therapy may enhance the therapeutic efficacy of doxorubicin in glioma cells by preventing EMT through inhibition of HDAC5. PMID: 26261519
- These results suggest a strong regulatory function of HDAC5 in the pro-inflammatory response of macrophages. PMID: 26059794
- In erythroid cells, pull down experiments identified the presence of a novel complex formed by HDAC5, GATA1, EKLF and pERK which was instead undetectable in cells of the megakaryocytic lineage. PMID: 24594363
- Data reveal a novel role of HDAC5 in modulating the KLF2 transcriptional activation and eNOS expression. PMID: 25096223
- Studied phosphorylation sites within functional HDAC5 domains, including the deacetylation domain (DAC, Ser755), nuclear export signal (NES, Ser1108), and an acidic domain (AD, Ser611). PMID: 24920159
- mRNA and protein levels of HDAC5 were up-regulated in human hepatocellular carcinoma. PMID: 25129440
- HDAC5 promoted the Six1 expression. PMID: 24706304
- In C2C12 myoblasts, recombinant human HDAC5 phosphorylation by PKD regulated the expression of diverse metabolic genes and glucose metabolism. PMID: 24732133
- findings show N-Myc upregulated HDAC5 expression in neuroblastoma cells; HDAC5 repressed NEDD4 gene expression,increased Aurora A gene expression and consequently upregulated N-Myc protein expression;data identify HDAC5 as a novel co-factor in N-Myc oncogenesis PMID: 23812427
- we show that Stat3 binds to the promoter region of PTPN13 and promotes its activity through recruiting HDAC5. Thus, our results suggest a previously unknown Stat3-PTPN13 molecular network controlling squamous cell lung carcinoma development PMID: 24191246
- At the molecular level, we demonstrated that HDAC5 promoted mRNA expression of twist 1, which has been reported as an oncogene PMID: 24092570
- These findings suggest that HDAC5 is a key determinant of p53-mediated cell fate decisions in response to genotoxic stress. PMID: 24120667
- Data indicate there was a link between baseline viral load, age (40 years), IL-28B (rs12979860), HDAC2 (rs3778216), HDAC3 (rs976552) and HDAC5 (rs368328) with sustained virological response (SVR). PMID: 23615070
- HDAC5 is essential for the length maintenance of long telomeres and its depletion is required for sensitization of cancer cells with long telomeres to chemotherapy. PMID: 23729589
- Loss of HDAC5 impairs memory function but has little impact in a transgenic mouse model of amyloid pathology. PMID: 22914591
- Nuclear calcium signaling is a regulator of nuclear export of HDAC4 and HDAC5. PMID: 23364788
- Dephosphorylation at a conserved SP motif governs cAMP sensitivity and nuclear localization of class IIa histone deacetylases HDAC4, 5 and 9 PMID: 23297420
- Data suggest that HDAC5 regulates muscle glucose metabolism and insulin action and that HDAC inhibitors can be used to modulate these parameters in muscle cells. PMID: 22991226
- The current study identified the class II deacetylase HDAC5 as a novel promoting factor of CTG*CAG expansions. PMID: 22941650
- HDAC5 in the maintenance/assembly of pericentric heterochromatin structure and demonstrate that class IIa HDAC5 can represent a potential target for anticancer therapies. PMID: 22301920
- The results of this study suggested that suggest that HDAC5 provides a delayed braking mechanism on gene expression programs that support the development, but not expression, of cocaine reward behaviors. PMID: 22243750
- Significantly increased methylation of the HDAC5 gene was associated with astrocytomas. PMID: 21508384
- Ser279 is a critical phosphorylation within the NLS involved in the nuclear import of HDAC5 PMID: 21081666
- in addition to activation of protein kinase D isozymes by phosphorylating Ser744 and Ser748 at their activation sites, PKCdelta may also play a role in the regulation of HDAC5 by phosphorylation of Ser259 PMID: 21146494
- differentiation-dependent GLUT4 gene expression in 3T3-L1 adipocytes is dependent on the nuclear concentration of a class II histone deacetylase (HDAC) protein, HDAC5 PMID: 21047791
- Findings identify HDAC5 as a substrate of PKA and reveal a cAMP/PKA-dependent pathway that controls HDAC5 nucleocytoplasmic shuttling and represses gene transcription. PMID: 20716686
- phosphorylation-dependent derepression of HDAC5 mediates flow-induced KLF2 and eNOS expression as well as flow anti-inflammation, and suggest that HDAC5 could be a potential therapeutic target for the prevention of atherosclerosis. PMID: 20042720
- Class II histone deacetylases are directly recruited by BCL6 transcriptional repressor PMID: 11929873
- Histone deacetylase 5 is not a p53 target gene, but its overexpression inhibits tumor cell growth and induces apoptosis. PMID: 12019172
- MITR, HDAC4, and HDAC5 associate with heterochromatin protein 1 (HP1), an adaptor protein that recognizes methylated lysines within histone tails and mediates transcriptional repression by recruiting histone methyltransferase PMID: 12242305
- HDAC5 binds to Ca(2+)/calmodulin and inhibits MEF2a binding PMID: 12626519
- ICP0 of herpes simplex virus Type 1 is able to overcome the HDAC5 amino-terminal- and MITR-induced MEF2A repression in gene reporter assays PMID: 15194749
- The HDAC5, a class II HDAC involved in myogenesis, was not detected in the tissues. PMID: 15590418
- G betagamma binds HDAC5 and inhibits its transcriptional co-repression activity PMID: 16221676
- novel transcriptional pathway under the control of class II HDACs and suggest a role for these transcriptional repressors as signal-responsive regulators of antigen presentation PMID: 16236793
- NO-dependent PP2A activation plays a key role in nuclear translocation of class II HDACs HDAC4 and HDAC5 PMID: 17975112
- AMP-activated protein kinase (AMPK) regulates GLUT4 transcription through the histone deacetylase (HDAC)5 transcriptional repressor. PMID: 18184930
顯示更多
收起更多
-
亞細(xì)胞定位:Nucleus. Cytoplasm. Note=Shuttles between the nucleus and the cytoplasm. In muscle cells, it shuttles into the cytoplasm during myocyte differentiation. The export to cytoplasm depends on the interaction with a 14-3-3 chaperone protein and is due to its phosphorylation at Ser-259 and Ser-498 by AMPK, CaMK1 and SIK1.
-
蛋白家族:Histone deacetylase family, HD type 2 subfamily
-
組織特異性:Ubiquitous.
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
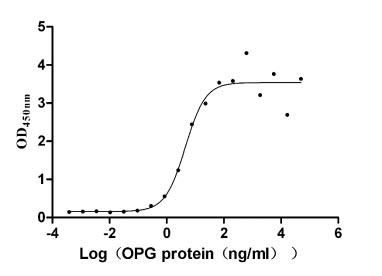
Recombinant Human Tumor necrosis factor receptor superfamily member 11B (TNFRSF11B) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
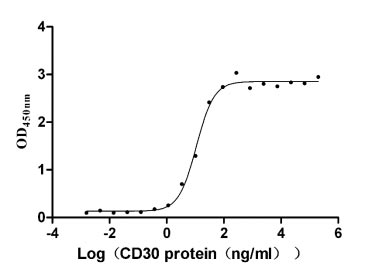
Recombinant Human Tumor necrosis factor ligand superfamily member 8 (TNFSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
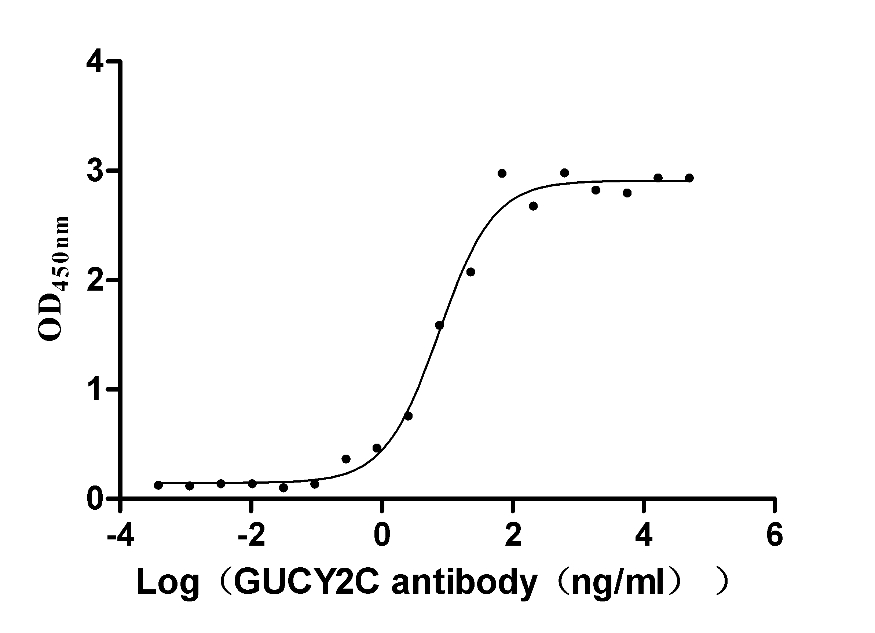
Recombinant Human Heat-stable enterotoxin receptor (GUCY2C), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
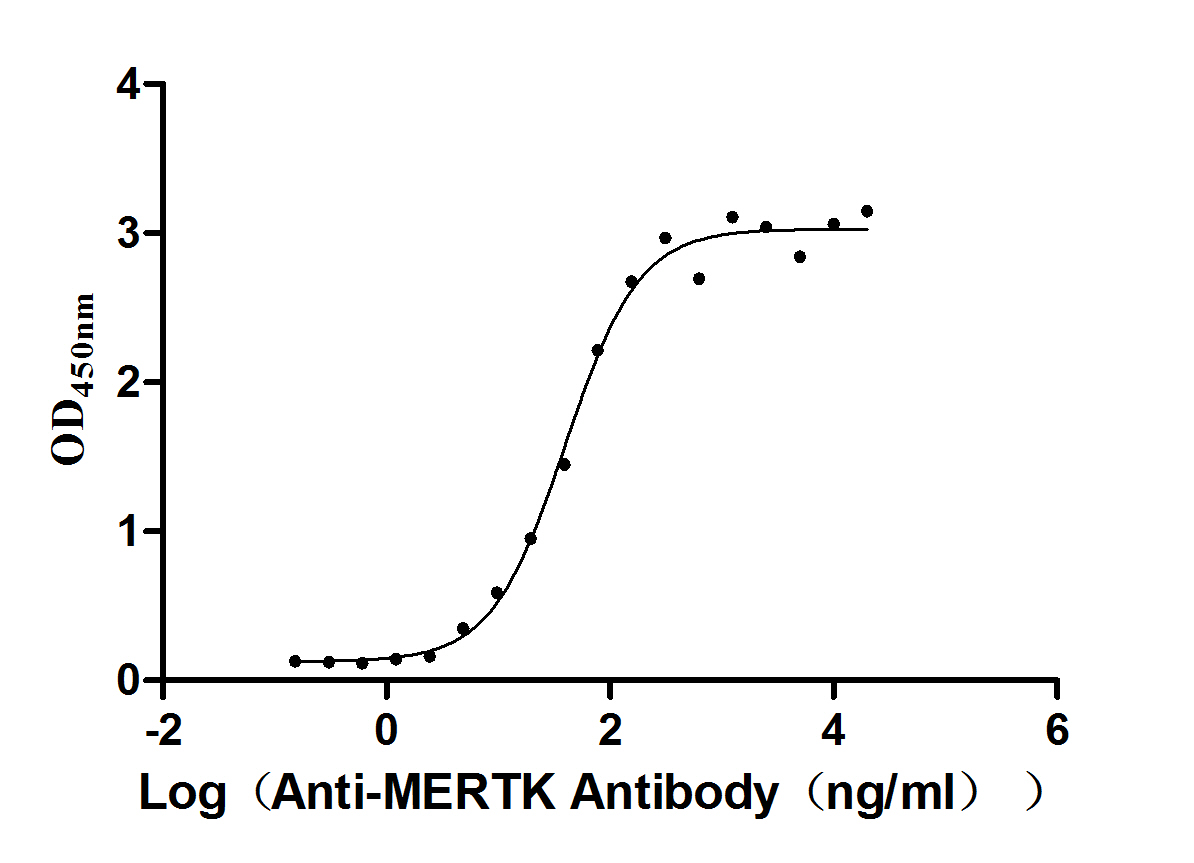
Recombinant Human Tyrosine-protein kinase Mer (MERTK), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
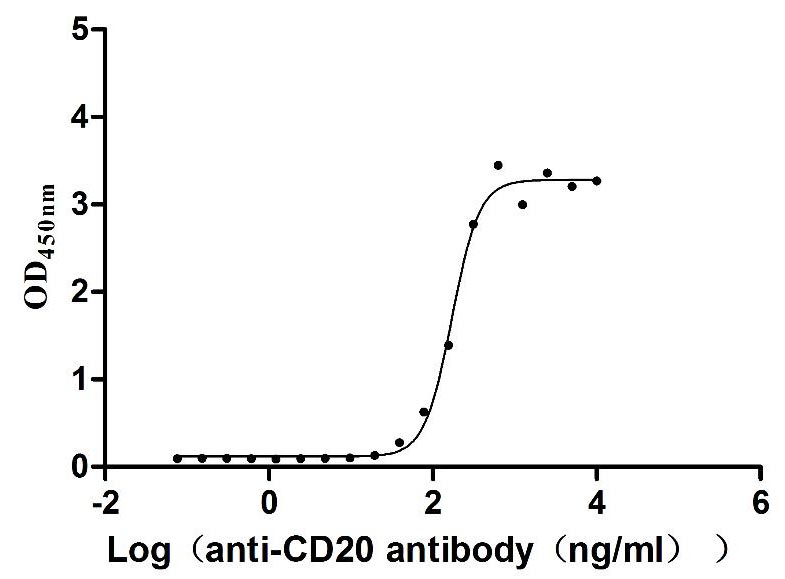
Recombinant Dog B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Canis lupus familiaris (Dog) (Canis familiaris)
-
-AC1.jpg)
Recombinant Human Claudin-6 (CLDN6)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
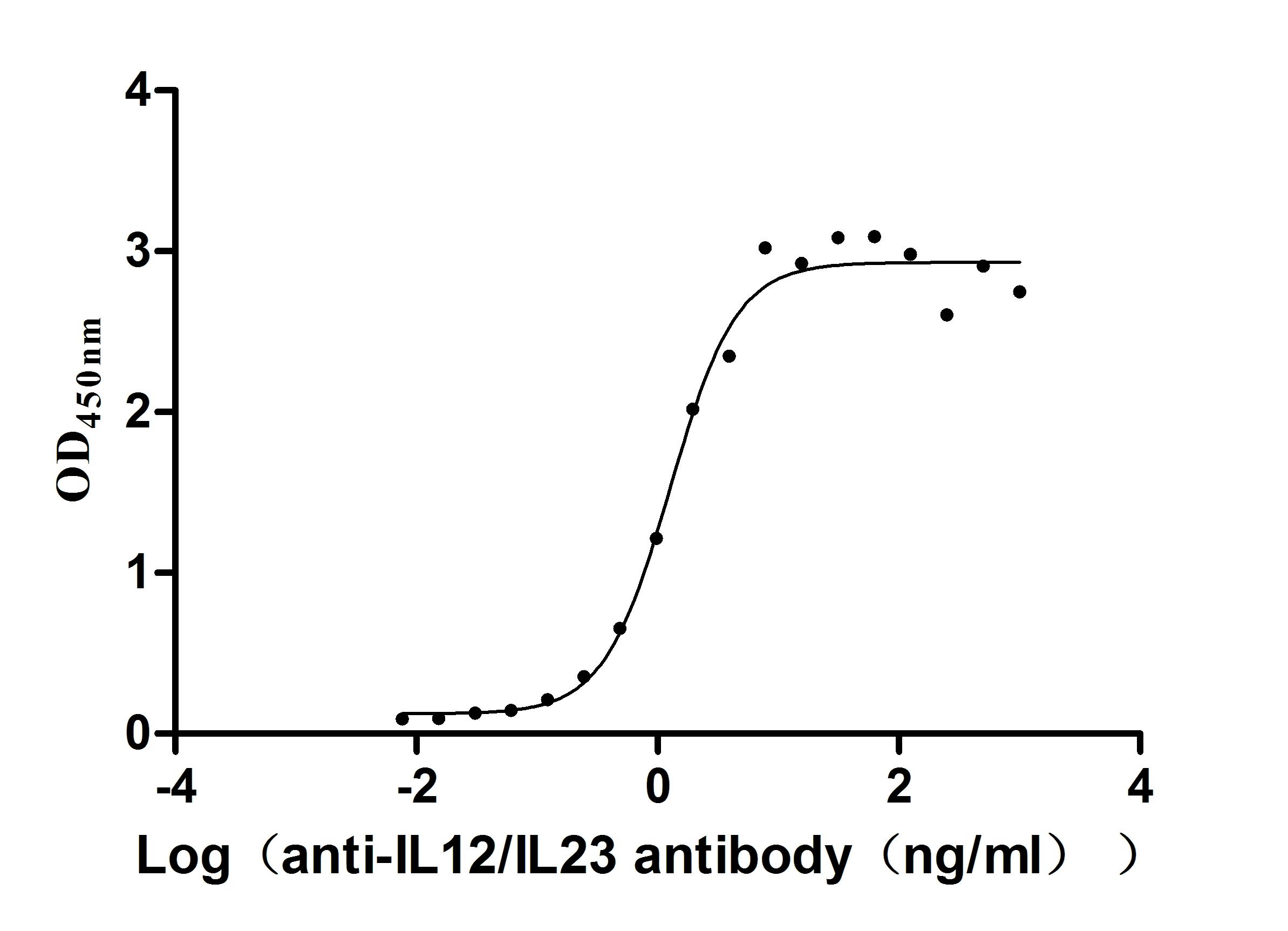
Recombinant Human IL12B&IL12A Heterodimer Protein (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
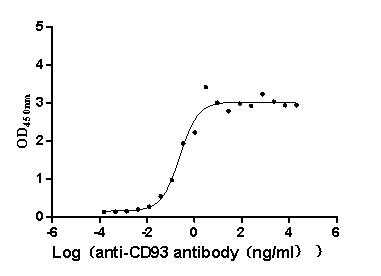
Recombinant Macaca fascicularis CD93 molecule (CD93), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)









