Recombinant Human Ataxin-3 (ATXN3)
In Stock-
中文名稱:Recombinant Human Ataxin-3 (ATXN3)
-
貨號:CSB-EP002443HU
-
規格:¥1836
-
圖片:
-
 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產品詳情
-
純度:Greater than 95% as determined by SDS-PAGE.
-
生物活性:Not Test
-
基因名:
-
Uniprot No.:
-
別名:Machado-Joseph disease protein 1;Spinocerebellar ataxia type 3 protein
-
種屬:Homo sapiens (Human)
-
蛋白長度:Full Length
-
來源:E.coli
-
分子量:48.7 kDa
-
表達區域:1-364aa
-
氨基酸序列MESIFHEKQEGSLCAQHCLNNLLQGEYFSPVELSSIAHQLDEEERMRMAEGGVTSEDYRTFLQQPSGNMDDSGFFSIQVISNALKVWGLELILFNSPEYQRLRIDPINERSFICNYKEHWFTVRKLGKQWFNLNSLLTGPELISDTYLALFLAQLQQEGYSIFVVKGDLPDCEADQLLQMIRVQQMHRPKLIGEELAQLKEQRVHKTDLERVLEANDGSGMLDEDEEDLQRALALSRQEIDMEDEEADLRRAIQLSMQGSSRNISQDMTQTSGTNLTSEELRKRREAYFEKQQQKQQQQQQQQQQGDLSGQSSHPCERPATSSGALGSDLGKACSPFIMFATFTLYLTYELHVIFALHYSSFPL
Note: The complete sequence may include tag sequence, target protein sequence, linker sequence and extra sequence that is translated with the protein sequence for the purpose(s) of secretion, stability, solubility, etc.
If the exact amino acid sequence of this recombinant protein is critical to your application, please explicitly request the full and complete sequence of this protein before ordering. -
蛋白標簽:C-terminal 6xHis-tagged
-
產品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:If the delivery form is liquid, the default storage buffer is Tris/PBS-based buffer, 5%-50% glycerol. If the delivery form is lyophilized powder, the buffer before lyophilization is Tris/PBS-based buffer, 6% Trehalose.
-
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:3-7 business days
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4℃ for up to one week.
-
Datasheet & COA:Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Deubiquitinating enzyme involved in protein homeostasis maintenance, transcription, cytoskeleton regulation, myogenesis and degradation of misfolded chaperone substrates. Binds long polyubiquitin chains and trims them, while it has weak or no activity against chains of 4 or less ubiquitins. Involved in degradation of misfolded chaperone substrates via its interaction with STUB1/CHIP: recruited to monoubiquitinated STUB1/CHIP, and restricts the length of ubiquitin chain attached to STUB1/CHIP substrates and preventing further chain extension. Interacts with key regulators of transcription and represses transcription: acts as a histone-binding protein that regulates transcription. Regulates autophagy via the deubiquitination of 'Lys-402' of BECN1 leading to the stabilization of BECN1.
-
基因功能參考文獻:
- Functionally, Ataxin-3 overexpression promoted cell proliferation, and Ataxin-3 knockdown inhibited cell proliferation in testicular cancer cell. In addition, up-regulation of Ataxin-3 inhibited the expression of PTEN and activated the AKT/mTOR pathway. PMID: 29902454
- Low exon skipping efficiencies combined with reduction in important ataxin-3 protein functions suggest that skipping of exon 8 and 9 is not a viable therapeutic option for spinocerebellar ataxia type-3. The modified protein was incapable of binding poly-ubiquitin chains, which may interfere with its normal deubiquitinating function. PMID: 27731380
- Data suggest ATXN3 binds with low-micromolar affinity to both wild-type p97/VCP and mutants linked to proteostasis deficiency multisystem proteinopathy 1 (MSP1; also called hereditary inclusion body myopathy); stoichiometry of binding is one ATXN3 molecule per p97/VCP hexamer in presence of ATP; MSP1 mutants of p97/VCP bind ATXN3 irrespective of nucleotide state. (VCP = valosin-containing protein/ATPase; ATXN3 = ataxin-3) PMID: 28939772
- DNA methylation levels in the ATXN3 promoter were significantly higher in SCA3/MJD patients PMID: 28094059
- data elucidate the important role of ataxin-3 proteolysis in the pathogenesis of Machado-Joseph disease. PMID: 28334907
- The findings reveal ATXN3 to be a novel deubiquitinase of Chk1, providing a new mechanism of Chk1 stabilization in genome integrity maintenance. PMID: 28180282
- Segregation patterns and factors influencing instability of expanded ATXN3 CAG transmissions in Machado-Joseph disease have been analyzed. PMID: 26693702
- Our data reveal a previously unrecognized balance between pathogenic and potentially therapeutic properties of the ataxin-3-Rad23 interaction; they highlight this interaction as critical for the toxicity of the SCA3 protein, and emphasize the importance of considering protein context when pursuing suppressive avenues. PMID: 28158474
- the opposing activities of RNF4 and ataxin-3 consolidate robust MDC1-dependent signaling and repair ofDNA double-strand break. PMID: 28275011
- we demonstrated that neural differentiation in these iPS cells was accompanied by autophagy and that rapamycin promoted autophagy through degradation of mutant ATXN3 proteins in neurally differentiated spinocerebellar ataxia-3 human induced pluripotent stem cells (p < 0.05). In conclusion, patient-derived iPS cells are a good model for studying the mechanisms of SCA3 and may provide a tool for drug discovery in vitro. PMID: 27847820
- South American cohort did not confirm the effect of the four candidate loci as modifier of onset age: mithocondrial A10398G polymorphism and CAGn at RAI1, CACNA1A, ATXN3, and ATXN7 genes PMID: 25869926
- Ataxin-3 phosphorylation decreases neuronal defects in spinocerebellar ataxia type 3 models. PMID: 26880203
- USP19_b up-regulates the protein levels of the polyglutamine (polyQ)-containing proteins, ataxin-3 (Atx3) and huntingtin (Htt), and thus promotes aggregation of their polyQ-expanded species in cell models PMID: 26808260
- Based on these data and other related studies, we presumed that de novo mutations of ATXN3 emerging from large ANs are at least one survival mechanisms of mutational ATXN3 and we can redefine the range of CAG repeats as: ANs/=50 PMID: 26266536
- Results suggest that the aggregation of Josephin proceeds from the monomer state to the formation of spheroidal intermediates with a native structure. Only successively, these intermediates evolve into misfolded aggregates and into the final fibrils. PMID: 26215704
- This study did not find relationship between CAG expansion length and psychiatric disorders. PMID: 26067219
- Data show that homozygosity for Machado-Joseph disease (MJD)/SCA3 protein enhances the clinical severity of the disease. PMID: 25566755
- A multistage aggregation mechanism for ataxin-3 is described in which flanking domain self-assembly precedes polyglutamine aggregation yet is influenced by polyglutamine expansion. PMID: 25700012
- Ubiquitination of ataxin-3 is not necessary for its proteasomal degradation.Ataxin-3 is regulated by ubiquitin-binding site 2 on its N terminus.Ubiquitin-binding site 2 of ataxin-3 prevents its proteasomal degradation by interacting with Rad23. PMID: 25144244
- Machado-Joseph disease patients carrying the rs709930 A allele and rs910369 T allele of ATXN3 experienced an earlier age of onset of approximately 2 to 4 years. PMID: 25689313
- polyglutamine expansion increases the molecular mobility of two juxtaposed helices critical to ataxin-3 deubiquitinase activity PMID: 26260925
- Evaluated the level of truncated pathological recombinant Ataxin-3 in a Drosophila model, in presence or absence of two suppressors and during aging. Suppressing truncated Ataxin-3-induced toxicity lowered the level of aggregated polyglutamine protein. PMID: 26210447
- Data support the importance of ATXN3 in neuronal cells and indicate that an expanded polyQ tract leads to a partial loss of the cellular function of ATXN3 that may be relevant to neurodegeneration. PMID: 25143392
- Wide type and polyQ-expanded ataxin-3 both showed partial co-localization to endoplasmic reticulum. PMID: 26037349
- Here we report that purified wild-type (WT) ATXN3 stimulates, and by contrast the mutant form specifically inhibits, PNKP's 3' phosphatase activity in vitro. ATXN3-deficient cells also show decreased PNKP activity PMID: 25633985
- We now report that the mutant ATXN3 protein interacts with and inactivates PNKP (polynucleotide kinase 3'-phosphatase), an essential DNA strand break repair enzyme PMID: 25590633
- The At3 N-terminal Josephin domain aggregation might be a multistep process. PMID: 25268243
- Results suggest that polyglutamine-expanded ataxin-3-Q79 impairs histone acetyltransferase activity, leading to impaired induction of cerebellar long-term depression in the spinocerebellar ataxia type 3 transgenic mouse PMID: 25139423
- Authors have investigated the interaction of AT3 with tubulin and HDAC6. PMID: 24685680
- miR-25 reduced both wild-type and polyQ-expanded mutant ataxin-3 protein levels by interacting with the 3'UTR of ATXN3 mRNA PMID: 25451224
- substrate recognition by the Josephin domain of ataxin-3 PMID: 25448680
- ataxin-3 fragment aggregates in a polyQ length-dependent manner in C. elegans muscle cells and that this aggregation is associated with cellular dysfunction PMID: 24817148
- In ATXN3-depleted cells, under conditions of transcriptional inhibition, PTEN and PTENP1 mRNAs rapidly decay. PMID: 24292675
- interaction between UbD2 and p97/Atx3 mediates retranslocation of UbD2 to the cytoplasm for terminal degradation in the proteasomes. PMID: 24196352
- Our work also suggests that ataxin-3 suppresses degeneration by regulating toxic protein aggregation rather than stability. PMID: 24106274
- The cloned A3IP gene encodes A3IP, a novel ataxin-3 interacting protein. PMID: 23926002
- Our data suggest that ataxin-3 plays an important role in regulating the Bcl-XL-Bax-mediated anti-oxidative response by modulating the interaction between Bcl-XL and Bax. PMID: 23562578
- This study demonistrated that Altered expression of carbonic anhydrase-related protein XI in neuronal cells expressing mutant ataxin-3. PMID: 23184527
- our study demonstrated that SUMOylation on K166, the first described residue of SUMO-1 modification of ataxin-3, partially increased the stability of mutant-type ataxin-3, and the rate of apoptosis arisen from the cytotoxicity of the modified protein PMID: 23382880
- 10 SNPs (none in core splicing signals) were found from exonic & flanking intronic regions in genomic DNA from Machado-Joseph disease patients & controls. The SNPs implied losses & gains of splicing-factor-recognition motifs. PMID: 22706685
- the efficacy of gene silencing in blocking the MJD-associated motor-behavior and neuropathological abnormalities PMID: 23349684
- Ataxin-3 is cleaved by calpains. Increased proteolytic cleavage of ataxin-3 results in a more severe and faster progressing neurological phenotype of spinocerebellar ataxia type 3. PMID: 23100324
- VCP/p97 was shown to be an activator specifically of wild-type ataxin-3. PMID: 22970133
- no functional effect could be predicted for ATXN3 gene variant. PMID: 22422287
- the sequestration of misfolded SOD1 into aggresomes, which is driven by ataxin-3, plays an important role in attenuating protein misfolding-induced cell toxicity. PMID: 22761419
- ATX3 proteolysis in transgenic mice by human calpains mediates ATX3 translocation to the nucleus, aggregation and toxicity. PMID: 22843411
- Transiently transfected HEK cell lines with expanded (Q84) ataxin-3 exhibited a higher susceptibility to 3-nitropropionic acid (3-NP), an irreversible inhibitor of mitochondrial complex II. PMID: 22037589
- These results underscore ataxin 3 capability of undergoing multiple aggregation pathways that lead to end products endowed with substantially different molecular structures. PMID: 22234302
- This study demonistrated that Disease progressed of autosomal dominant cerebellar ataxia and spastic paraplegia faster in SCA s with polyglutamine expansions in SCA1, 2, and 3. PMID: 22491195
- the central flexible region enhances protein aggregation and can populate conformational states with different degrees of compactness PMID: 22129356
顯示更多
收起更多
-
相關疾病:Spinocerebellar ataxia 3 (SCA3)
-
亞細胞定位:Nucleus matrix. Nucleus. Note=Predominantly nuclear, but not exclusively, inner nuclear matrix.
-
組織特異性:Ubiquitous.
-
數據庫鏈接:
Most popular with customers
-
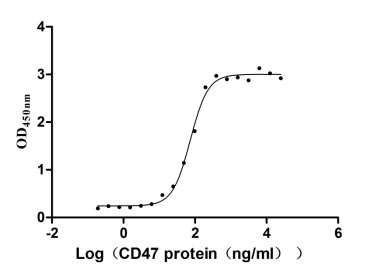
Recombinant Human Leukocyte surface antigen CD47 (CD47), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
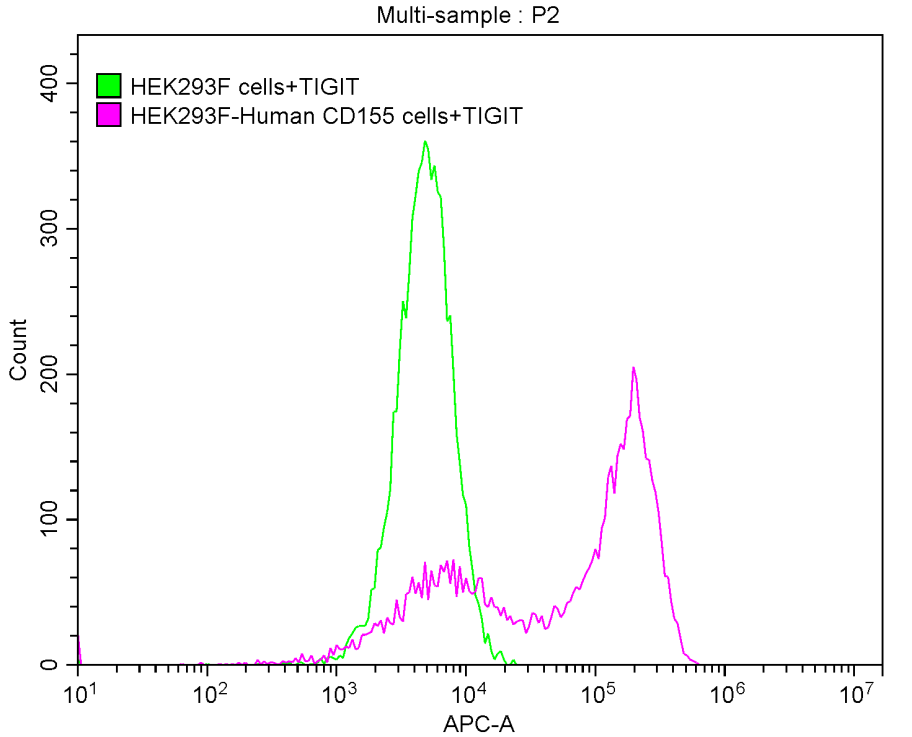
Recombinant Human T-cell immunoreceptor with Ig and ITIM domains (TIGIT), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
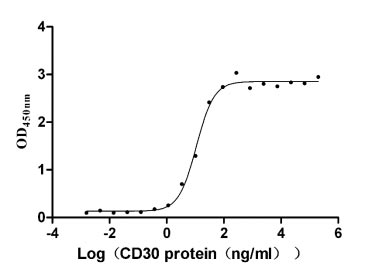
Recombinant Human Tumor necrosis factor ligand superfamily member 8 (TNFSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Human Claudin-18.2 (CLDN18.2)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
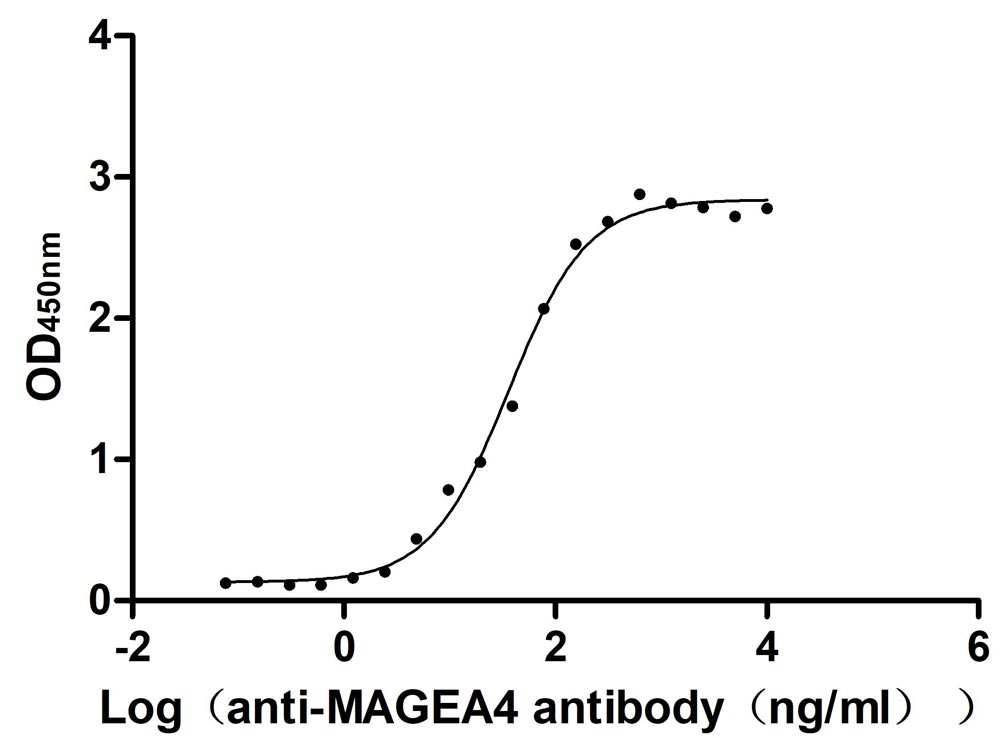
Recombinant Human Melanoma-associated antigen 4 (MAGEA4) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Mouse Retinol-binding protein 4 (Rbp4) (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
Recombinant Human Tumor-associated calcium signal transducer 2 (TACSTD2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human CUB domain-containing protein 1 (CDCP1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









