Recombinant Drosophila melanogaster Cryptochrome-1 (cry), partial
-
中文名稱:Recombinant Drosophila melanogaster Cryptochrome-1(cry) ,partial
-
貨號:CSB-YP006005DLU
-
說明書:
-
規格:
-
來源:Yeast
-
其他:
-
中文名稱:Recombinant Drosophila melanogaster Cryptochrome-1(cry) ,partial
-
貨號:CSB-EP006005DLU
-
說明書:
-
規格:
-
來源:E.coli
-
其他:
-
中文名稱:Recombinant Drosophila melanogaster Cryptochrome-1(cry) ,partial
-
貨號:CSB-EP006005DLU-B
-
說明書:
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Drosophila melanogaster Cryptochrome-1(cry) ,partial
-
貨號:CSB-BP006005DLU
-
說明書:
-
規格:
-
來源:Baculovirus
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:cry
-
Uniprot No.:
-
別名:cry; CG3772Cryptochrome-1; DmCRY1; dcry; Blue light photoreceptor
-
種屬:Drosophila melanogaster (Fruit fly)
-
蛋白長度:Partial
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder Warning: in_array() expects parameter 2 to be array, null given in /www/web/cusabio_cn/public_html/caches/caches_template/default/content/show_product_protein.php on line 662
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Blue light-dependent regulator that is the input of the circadian feedback loop. Has no photolyase activity for cyclobutane pyrimidine dimers or 6-4 photoproducts. Regulation of expression by light suggests a role in photoreception for locomotor activity rhythms. Functions, together with per, as a transcriptional repressor required for the oscillation of peripheral circadian clocks and for the correct specification of clock cells. Genes directly activated by the transcription factors Clock (Clk) and cycle (cyc) are repressed by cry. Necessary for light-dependent magnetosensitivity, an intact circadian system is not required for the magnetoreception mechanism to operate. Required for both the naive and trained responses to magnetic field, consistent with the notion that cry is in the input pathway of magnetic sensing.
-
基因功能參考文獻:
- photo-induced electron transfer reactions in Drosophila melanogaster cryptochrome are indeed influenced by magnetic fields of a few millitesla. PMID: 28176875
- Substitutions of four key Trp residues to redox-active tyrosine and redox-inactive phenylalanine tune the light sensitivity of dCRY photoreduction, conformational activation, cellular stability, and targeted degradation of the clock protein timeless PMID: 29581265
- we identify the circadian blue-light photoreceptor CRYPTOCHROME as a molecular regulator of Prolonged Morning Wakefulness (PMW), and propose a model in which the Drosophila nervous system integrates information encoding temperature, light, and time to dynamically control when sleep is initiated. Our results provide a platform to investigate how environmental inputs co-ordinately regulate sleep plasticity. PMID: 28084307
- The findings of this article for the first time define CRY expression in Drosophila peripheral tissues and reveal that CRY acts together with K(+) channels to maintain passive membrane properties in a non-clock-containing peripheral tissue independent of light. PMID: 28781048
- Changes in active site histidine hydrogen bonding trigger cryptochrome activation. PMID: 27551082
- CRY is not stabilized by interaction with the kinase Shaggy (SGG), the GSK-3 beta fly orthologue. PMID: 26741981
- our studies provided novel evidence that the circadian clock gene, dCry, plays an essential role in heart morphogenesis and function. PMID: 26348211
- Using time-resolved and steady-state optical spectroscopy, we studied the mechanism of light-induced radical-pair formation and decay, and the photoreduction of the FAD cofactor. PMID: 25879256
- The results of this study concluded that the E oscillators are the targets of light input via CRY and the visual system and are required for normal light entrainment. PMID: 25878285
- The Cry leucine-histidine substitution is common in Drosophila, with both alleles at intermediate frequencies across 27 populations surveyed in Europe, irrespective of latitude. PMID: 24475129
- Both oxidized and reduced forms of dCRY are capable of photosignaling. PMID: 24379403
- Flavin reduction activates Drosophila cryptochrome. PMID: 24297896
- Our findings indicate that CRY compensates for limited light sensitivity in vivo by photon integration over extraordinarily long times, and point to select circadian pacemaker neurons as having important roles. PMID: 23874218
- dampens temperature-induced PERIOD oscillations in dorsal clock neurons PMID: 23333312
- Study reports crystal structures of full-length dCRY, a dCRY loop deletion construct, and the photolyase homology region of mouse CRY1 (mCRY1). PMID: 23746849
- Data indicate that wild-type flies show greatly enhanced magnitude of phase shift, but cryptochrome (CRY)-less flies seem impaired in the ability to integrate duration of the light pulse in a wild-type manner. PMID: 22476772
- Study reports that increased nighttime activity of Clk mutants is mediated by high levels of the circadian photoreceptor CRYPTOCHROME (CRY) in large ventral lateral neurons (l-LN(v)s). PMID: 22581798
- Drosophila photolyases contain F0 as the second chromophore; within the limitation of the current assay, neither plant nor animal cryptochromes seem to carry a second chromophore. PMID: 22175817
- 2.3-A resolution crystal structure of Drosophila CRY with an intact C terminus PMID: 22080955
- Data show that CRY is a key component for the preferential entrainment to light. PMID: 21135155
- study shows the CRY-mediated light response requires a flavin redox-based mechanism and depends on potassium channel conductance but is independent of the classical circadian CRY-TIMELESS interaction PMID: 21385718
- CRY's photolyase homology domain is sufficient for light detection and phototransduction, whereas the carboxyl-terminal domain regulates CRY stability, CRY-TIM interaction, and circadian photosensitivity PMID: 15178801
- Cryb mutant flies exhibited free-running rhythms with two rhythmic components. Drosophila locomotor rhythms may be driven by two separate PER-dependent clocks, responding differentially to constant light. PMID: 15183277
- show a strong interaction between Veela and cryptochrome genetic variants, demonstrating that the Jetlag, Timeless, and Cry proteins function in the same pathway PMID: 17068124
- Cryptochrome is present in the compound eyes and a subset of Drosophila's clock neurons PMID: 18399544
- CRY is not a core component, but an exclusive photoreceptor in the cuticle deposition rhythm. PMID: 18539772
- the first genetic evidence for a Cry-based magnetosensitive system in any animal PMID: 18641630
- Spatial and cricadian regulation of cry in Drosophila are reported. PMID: 18663236
- cry is expressed in a subset of circadian oscillator neurons in the Drosophila CNS. PMID: 18663237
- After illumination, Jetlag protein induces massive degradation of Cry as part of the resetting process of the circadian clock in reaction to light. PMID: 19185492
- These results suggest that cry, Rh1, Rh5 and Rh6 are essential for circadian photoentrainment to green and yellow light. PMID: 19398933
顯示更多
收起更多
-
亞細胞定位:Cytoplasm. Cytoplasm, perinuclear region. Nucleus.
-
蛋白家族:DNA photolyase class-1 family
-
組織特異性:Expressed at higher levels in the head than in body and it is more expressed in antennae than in legs, wings and mouth appendages. Prominent expression is seen in cells of the lateral brain, which are close to or coincident with the clock neurons. Abundan
-
數據庫鏈接:
Most popular with customers
-
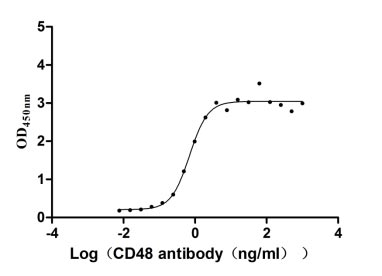
Recombinant Human CD48 antigen (CD48) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
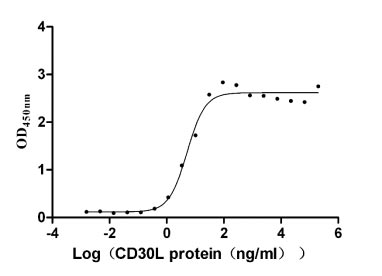
Recombinant Human Tumor necrosis factor ligand superfamily member 8 (TNFSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
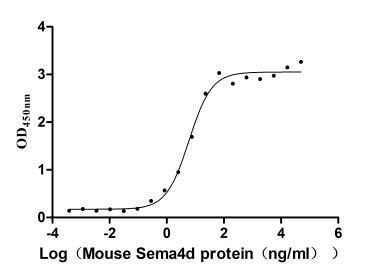
Recombinant Mouse Semaphorin-4D (Sema4d), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
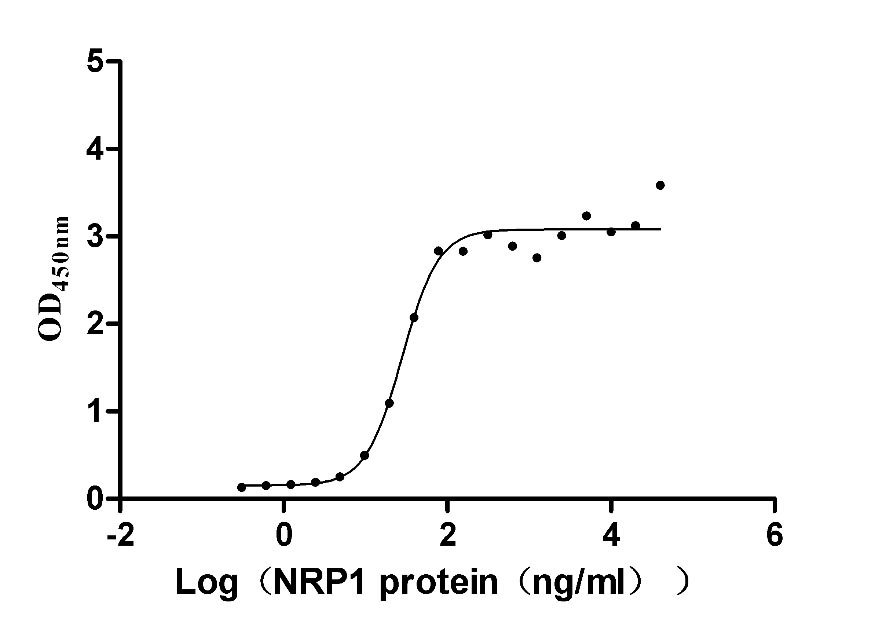
Recombinant Human Neuropilin-1 (NRP1) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Dog B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Canis lupus familiaris (Dog) (Canis familiaris)
-
Recombinant Human Desmoglein-2 (DSG2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Serine/threonine-protein kinase receptor R3 (ACVRL1), partial (Active)
Express system: Baculovirus
Species: Homo sapiens (Human)
-
Recombinant Human C-C chemokine receptor type 6(CCR6)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









