造血干細胞表面標志物
日期:2020-01-10 13:43:12
造血干細胞(Hematopoietic stem cells, HSC)是一類成體干細胞,具有干細胞的特性:自我更新和分化潛能。
造血干細胞是一種骨髓來源的多能干細胞,它是血液系統中的“種子”細胞。造血干細胞(HSCs)在造血過程中形成血系中的所有細胞,包括各種成熟細胞如白細胞、紅細胞、血小板等。在受到適當刺激時還分化為其他非造血組織(如脂肪細胞、心肌細胞、內皮細胞和胰腺細胞)[1]。
造血干細胞具有調節體內平衡、免疫功能、抗微生物、抗炎癥等生物學功能。它在血液病、遺傳性血液病和自身免疫性疾病的治療方面有重要作用。
1. 造血干細胞類型
已經定義了兩種類型的造血干細胞:能夠終生保持自我更新和多譜系分化潛能的長期再生細胞(LTRC);來源于LTRC的短期再生細胞(STRC),雖然它們保持了多能性,但它們表現出更有限的自我更新潛能。它們重建髓系和/或淋巴系間隔的時間很短,大約6周。
2. 造血干細胞的來源
造血干細胞存在于成年人的骨髓中,特別是在骨盆、股骨和胸骨中。它們也存在于臍帶血和少量的外周血中。
2.1 骨髓
造血干細胞(HSCs)是一種骨髓來源的多能干細胞。
從骨髓中獲取造血干細胞通過外科手術,從兩個髂骨后嵴分次采集。骨髓中每10萬個細胞中約有1個是長期造血干細胞(LT-HSC)。
2.2 外周血
大部分造血干細胞來源于骨髓,少量的干細胞和祖細胞在血液中循環。人類造血干細胞的臨床移植,可以從外周血中收集供體細胞。造血干細胞的采集是在粒細胞集落刺激因子(G-CSF)等造血生長因子的作用下,將骨髓中的HSC動員到外周血后,通過分離的方式采集。
2.3 臍帶血(UCB)
臍帶血是造血干細胞和造血祖細胞的豐富來源,它所含不同類型的造血祖細胞的數量大約是成人血液中觀察到的數量的10倍。臍帶血作為再生醫學的特殊細胞來源,也含有多種類型的干細胞。
2.4 胎兒造血系統與胚胎造血干細胞
胎兒造血系統是造血干細胞的重要來源,但在臨床上尚未得到應用。
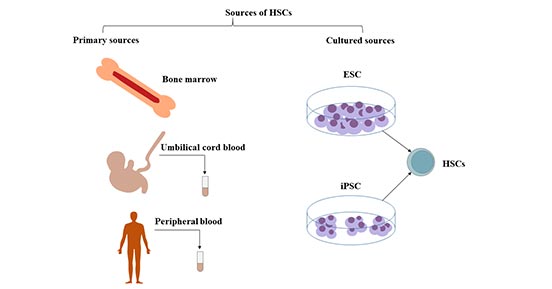
圖1. 造血干細胞的來源
3. 造血干細胞的生物學特性
造血干細胞是真正的干細胞,因為它們具有多能性、自我更新能力。其他特性還包括它的異質性。
自我更新:造血干細胞的自我更新意味著造血干細胞產生與它們相同的子代細胞。它的這種能力,使得造血干細胞可以從單個細胞產生一個完整的造血系統,并在個體的一生中保持造血。
多能性:是指造血干細胞在需要時產生主要造血細胞類型的能力。造血干細胞(HSC)可分化為來自髓系(單核細胞和巨噬細胞、中性粒細胞、嗜堿性粒細胞、嗜酸性粒細胞、紅細胞、巨核細胞/血小板、樹突狀細胞)和淋巴系(T、B和NK細胞)的各種血細胞。
造血干細胞具有極高的增殖潛能,使其能夠滿足正常成人一生中對造血的高需求。
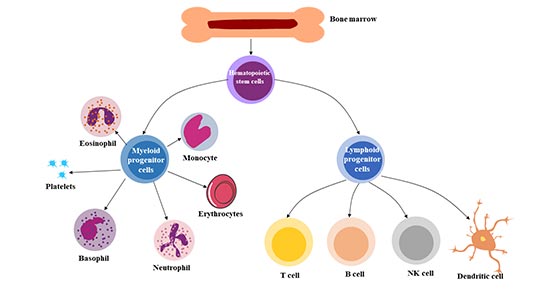
圖2. 造血干細胞的多能性
可塑性:可塑性是指造血干細胞分化為多種非造血組織如心肌細胞的能力[2]。
造血干細胞運輸:從胚胎起源開始,HSPC從一個生態位移動到另一個。造血干細胞(HSC)生態位是造血干細胞賴以生存和自我更新的空間位置和生理微環境。造血干細胞的運輸可分為歸巢、保留和植入。
異質性:造血干細胞內部的異質性是指造血干細胞具有不同的生理特征,如細胞周期狀態[3]和自我更新能力[4],對不同的外部信號有不同的反應,在移植后有不同的譜系輸出[5][6][7][8]。
造血干細胞內部的異質性是構成成人造血系統基礎的最堅實的細胞部分,為什么存在HSC亞型、它們是如何產生的?
Muller-Sieberg組是最早確定HSC異質性的研究小組之一。HSC的異質性可以穩定地傳播。HSC的異質性是如何產生的呢?目前認為造血干細胞(HSC)異質性產生的可能途徑如下:
- 造血干細胞來源不同:HSC類型可能來自中胚層細胞、內皮細胞或來自于背主動脈前造血干細胞。
- 調控因素的差異:HSC的類型可能受不同的誘導性發育組織(如AGM、胎盤、卵黃囊、頭部)的調節,并可能隨著它們在循環中的遷移而改變;或受發育生態位(可能是血管、肝臟、神經或骨骼)的調節;或受不同的發育機制(如EHT)的調節。
- 此外,表觀遺傳修飾也可能解釋不同HSC類型的存在。
4. HSC和免疫系統
造血干細胞(HSC)生態位是造血干細胞賴以生存和自我更新的空間位置和生理微環境。它保持干細胞在自我更新和分化之間的動態平衡。
研究表明,在活體內,造血干細胞的生態位為造血干細胞提供了免疫特權部位。調節性T細胞(Treg)與造血干細胞定位于骨髓的骨內膜區域,以保護造血干細胞免受免疫攻擊。
此外,造血干細胞本身也通過調節表面免疫分子的表達而具有一定的免疫豁免潛能。
當HSCs被強炎癥信號激活并動員進入循環時,HSCs表面的CD47水平顯著上調。CD47與巨噬細胞上的信號調節蛋白結合,抑制吞噬作用。CD47在動員的造血干細胞表面表達的增加可以保護這些細胞不被吞噬。
5. 人體造血系統層次
造血系統中所有血細胞的起源被認為來自于具有自我更新能力的造血干細胞。
造血干細胞和祖細胞(HSPCs)庫可分為三種類型的細胞:長期造血干細胞(LT-HSC)、短期造血干細胞(ST-HSPC)和多能祖細胞(MPP)。
造血系統中,靜止的、長期再填充的造血干細胞(LT-HSC)位于所有成熟血細胞的頂端,通過其自我更新和不對稱的細胞分裂潛能在一生中維持一個原始的多潛能池。在穩定狀態下,造血干細胞衍生的子代具有對外周刺激的快速增殖能力,取代了活化的、消耗的或老化的血細胞的損失[9][10]。
ST-HSPC及其祖細胞具有維持正常造血6~8周的能力。喪失正常的LT-HSC和ST-HSPC功能是自然干細胞老化和幾種造血疾病的標志,最顯著的是血液系統惡性腫瘤的發展和進展[11]。
多能祖細胞(MPPs)是由造血干細胞產生的失去自我更新潛能但仍能完全分化為所有多系的多潛能祖細胞(MMPPs)。MPPs進一步產生低潛能祖細胞,即常見的淋巴和髓系祖細胞(CLPs和CMPs)。
所有這些低潛能祖細胞均可分化為有限的定型譜系:
CMPs可分化為巨核/紅細胞祖細胞(MEPs)、粒/巨噬細胞祖細胞(GMPs)和樹突狀細胞(DC)祖細胞;
CLPs可分化為T細胞祖細胞、B細胞祖細胞、NK細胞祖細胞和DC祖細胞。[12][13]。
DC祖細胞(CD8+DC、CD8-DC和漿細胞樣DC)可以來自CMPs和CLPs。
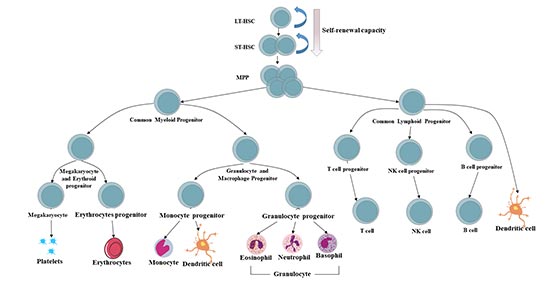
圖3. 人體造血系統層次
6. 造血干細胞的細胞表面標志
盡管關于造血干細胞的研究很多,但是還沒有發現一個單獨的分子標記是由造血干細胞表達。由于造血干細胞缺乏成熟血細胞標志物的表達,因此被稱為Lin-。對造血干細胞進行鑒定或分離一般都是采用多個不同的細胞表面標志物的組合,利用流式細胞術將稀少的造血干細胞從周圍的血細胞中分離出來。
人造血干細胞與小鼠造血干細胞的標記物在普遍接受的造血干細胞類型上存在許多差異。
小鼠HSC:EMCN+、CD34lo/-、SCA-1+、Thy1.1+/lo、CD38+、C-kit+、lin-
人HSC:EMCN+、CD34+、CD59+、Thy1/CD90+、CD38lo/-、C-kit/CD117+、lin-
區分鼠標長期(LT-HSC)和短期(ST-HSC)造血干細胞和多能祖細胞(早期MPP和晚期MPP)標記如下:
LT-HSC:CD34-、CD38-、SCA-1+、Thy1.1+/lo、C-kit+、lin-、CD135-、Slamf1/CD150+
ST-HSC:CD34+、CD38+、SCA-1+、Thy1.1+/lo、C-kit+、lin-、CD135-、Slamf1/CD150+、Mac-1 (CD11b)lo
早期MPP:CD34+、SCA-1+、Thy1.1-、C-kit+、lin-、CD135+、Slamf1/CD150-、Mac-1 (CD11b)lo、CD4lo
晚期MPP:CD34+、SCA-1+、Thy1.1-、C-kit+、lin-、CD135high、Slamf1/CD150-、Mac-1 (CD11b)lo、CD4lo
6.1 主要標志物的介紹
6.1.1 CD34
CD34是最重要的標志之一,在造血干細胞及其祖細胞的分離和鑒定中,CD34分子是第一個被廣泛研究的分子。CD34在人臍帶血、骨髓和外周血中的表達約0.1 - 4.9%[14]。CD34在0.55%的人骨髓細胞上表達,在早期祖細胞上有表達,而在成熟的骨髓細胞上不表達。其他表面標記已經與CD34結合使用來區分原始細胞群體。
6.1.2 CD38
CD38又稱環狀ADP核糖水解酶,是一種存在于許多免疫細胞(白細胞)表面的糖蛋白,包括CD4+、CD8+、B淋巴細胞和自然殺傷細胞。CD38標記用于區分造血干細胞多能祖細胞(CD38-)和定向祖細胞(CD38+)。
6.1.3 CD90 (Thy1)
CD90,也被稱為Thy-1,是一個28-30 kDa 的GPI連接膜糖蛋白。CD90表達于造血干細胞、神經元、胸腺細胞、外周T細胞、成纖維細胞和基質細胞。
CD90在CD34 + CD38-上的共表達定義了造血干細胞,而CD34 + CD38- CD90-定義了多能祖細胞。
6.1.4 CD117 (C-Kit)
CD117是一種145 kDa的蛋白酪氨酸激酶,也被稱為c-Kit。CD117在多能造血祖細胞(約1-4%的骨髓細胞)、肥大細胞和急性髓系白血病細胞(AML)上表達。
通過與它的配體結合可誘導CD117的磷酸化,刺激原始造血干細胞、紅細胞和單核細胞的增殖和存活。在配子形成、黑色素形成和造血過程中起著重要作用。
6.1.5 CD135 (Flk-2)
CD135,又稱FLK-2、FLT3和Ly-72,是一種III型酪氨酸激酶受體。CD135既表達在正常的CD34+造血干細胞中,也表達于惡性造血細胞,包括AML、ALL和CML BC。
CD135與FLT3配體結合,可以調節造血干細胞的生長,促進具有髓系和B淋巴系潛能的原始造血祖細胞的存活。
6.1.6 CD150 (SLAM)
CD150又稱SLAM,I型跨膜糖蛋白信號轉導淋巴細胞活化分子,是CD2蛋白家族SLAM亞群的典型成員。
SLAM表達于胸腺細胞、T細胞亞群、B細胞、樹突狀細胞、巨噬細胞和造血干細胞。
6.1.7 CD184 (CXCR4)
CD184,又稱Fusin或CXCR4。它廣泛表達于血液和組織細胞,包括B細胞和T細胞、單核細胞、巨噬細胞、樹突狀細胞、粒細胞、巨核細胞/血小板、淋巴樣細胞、髓樣前體細胞、內皮細胞、上皮細胞、星形膠質細胞和神經元等組織細胞。
在骨髓龕中,造血干細胞表達CXCR4。CXCR4是趨化因子CXCL12 (SDF-1)的受體,介導血細胞遷移,參與B淋巴細胞和骨髓生成、心臟發生、血管形成和小腦發育。
CXCR4和SDF-1之間的相互作用負責HSC在小生境中的歸巢和保留。HSC生態位中CXCR4/CXCL12 (SDF-1)的相互作用被認為是HSC動員的機制[15]。
6.1.8 Ly-6A/E (Sca-1)
Ly-6A/E,又稱Sca-1,是Ly-6多基因家族的成員,是一種表達于造血干細胞上的糖基磷脂酰肌醇(GPI)連接蛋白。其在多能造血干細胞(HSC)上的表達已被用作兩種Ly 6單倍體小鼠HSC的標記物。在成年動物的骨髓、胎肝、動員后的外周血和脾臟中均可見Sca-1陽性的造血干細胞。Ly-6A/E被認為參與了T細胞和B細胞反應的調節。
7. HSC的臨床應用
7.1 造血細胞移植(HCT)
造血細胞移植按照干細胞來源可分為骨髓移植、外周血干細胞移植和臍血移植等。
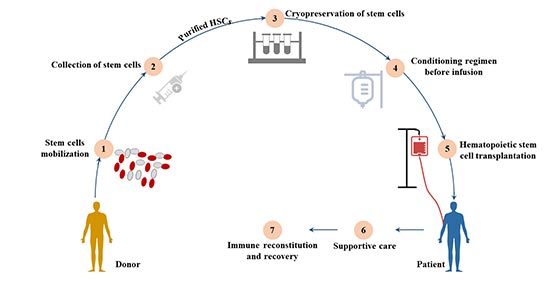
圖4. 造血細胞移植
7.1.1 骨髓移植
骨髓移植是造血干細胞移植的一種常見類型,造血干細胞在骨髓移植中對患者血細胞的再增殖起著至關重要的作用。
按其供受者關系大體可分為自體造血干細胞移植和異體造血干細胞移植(包括同胞間、非親緣間移植)。除了實體瘤外,骨髓移植還被用于治療白血病和免疫系統紊亂等血液疾病[16]。
7.1.2 外圍造血干細胞移植
外周血中循環的HSC在給予GM-CSF、IL-3或SCF等造血細胞因子后,其濃度可提高100倍。使用細胞分離器,通常可以從外周血中獲得所需數量的HSC。
7.1.3 臍帶血移植(UCB)
臍帶血容易獲得,且對捐贈者沒有風險,因此是一個有吸引力的可移植造血干細胞的來源。
新生兒免疫系統的不成熟,臍帶血中存在的淋巴細胞已被證明對HLA錯配有更強的耐受性,因此在異基因移植環境中引發的急性和慢性移植物抗宿主病(GVHD)較少。
7.2 其他
造血干細胞還在包括移植物抗腫瘤治療、耐受誘導、基因治療[17]、再生醫學等方面有廣泛的應用。
References
[1] Chotinantakul K, Leeanansaksiri W. Hematopoietic stem cell development, niches, and signaling pathways [J]. Bone marrow research, 2012, 2012.
[2] Chatterjee T, Sarkar R S, Dhot P S, et al. Adult stem cell plasticity: Dream or reality? [J]. Medical journal, Armed Forces India, 2010, 66(1): 56.
[3] Wilson A, Laurenti E, Oser G, et al. Hematopoietic stem cells reversibly switch from dormancy to self-renewal during homeostasis and repair [J]. Cell, 2008, 135(6): 1118-1129.
[4] Ema H, Morita Y, Suda T. Heterogeneity and hierarchy of hematopoietic stem cells [J]. Experimental hematology, 2014, 42(2): 74-82. e2.
[5] Benz C, Copley M R, Kent D G, et al. Hematopoietic stem cell subtypes expand differentially during development and display distinct lymphopoietic programs [J]. Cell stem cell, 2012, 10(3): 273-283.
[6] Dykstra B, Kent D, Bowie M, et al. Long-term propagation of distinct hematopoietic differentiation programs in vivo [J]. Cell stem cell, 2007, 1(2): 218-229.
[7] Sieburg H B, Cho R H, Dykstra B, et al. The hematopoietic stem compartment consists of a limited number of discrete stem cell subsets [J]. Blood, 2006, 107(6): 2311-2316.
[8] Verovskaya E, Broekhuis M J C, Zwart E, et al. Heterogeneity of young and aged murine hematopoietic stem cells revealed by quantitative clonal analysis using cellular barcoding [J]. Blood, 2013, 122(4): 523-532.
[9] Busch K, Klapproth K, Barile M, et al. Fundamental properties of unperturbed haematopoiesis from stem cells in vivo [J]. Nature, 2015, 518(7540): 542.
[10] Sawai C M, Babovic S, Upadhaya S, et al. Hematopoietic stem cells are the major source of multilineage hematopoiesis in adult animals [J]. Immunity, 2016, 45(3): 597-609.
[1] Sun J, Ramos A, Chapman B, et al. Clonal dynamics of native haematopoiesis [J]. Nature, 2014, 514(7522): 322.
[11] Yao J C, Link D C. Concise review: the malignant hematopoietic stem cell niche [J]. Stem Cells, 2017, 35(1): 3-8.
[12] Adams G B, Scadden D T. The hematopoietic stem cell in its place [J]. Nature immunology, 2006, 7(4): 333.
[13] Mosaad Youssef Mohamed,Hematopoietic stem cells: an overview. [J] .Transfus. Apher. Sci., 2014, 51: 68-82.
[14] Pranke P, Hendrikx J, Debnath G, et al. Immunophenotype of hematopoietic stem cells from placental/umbilical cord blood after culture [J]. Brazilian journal of medical and biological research, 2005, 38(12): 1775-1789.
[15] Herbert K E, Levesque J P, Mills A K, et al. How we mobilize haemopoietic stem cells [J]. Internal medicine journal, 2011, 41(8): 588-594.
[16] Rogers I, Casper R F. Umbilical cord blood stem cells [J]. Best Practice & Research Clinical Obstetrics & Gynaecology, 2004, 18(6): 893-908.
[17] Hirschi K K. Hemogenic endothelium during development and beyond [J]. Blood, 2012, 119(21): 4823-4827.
上一篇: 間充質干細胞的表面標記物









