Recombinant Rat Circadian locomoter output cycles protein kaput (Clock), partial
-
中文名稱:Recombinant Rat Circadian locomoter output cycles protein kaput(Clock) ,partial
-
貨號:CSB-YP894389RA
-
說明書:
-
規格:
-
來源:Yeast
-
其他:
-
中文名稱:Recombinant Rat Circadian locomoter output cycles protein kaput(Clock) ,partial
-
貨號:CSB-EP894389RA
-
說明書:
-
規格:
-
來源:E.coli
-
其他:
-
中文名稱:Recombinant Rat Circadian locomoter output cycles protein kaput(Clock) ,partial
-
貨號:CSB-EP894389RA-B
-
說明書:
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Rat Circadian locomoter output cycles protein kaput(Clock) ,partial
-
貨號:CSB-BP894389RA
-
說明書:
-
規格:
-
來源:Baculovirus
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:ClockCircadian locomoter output cycles protein kaput; rCLOCK; EC 2.3.1.48
-
種屬:Rattus norvegicus (Rat)
-
蛋白長度:Partial
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Transcriptional activator which forms a core component of the circadian clock. The circadian clock, an internal time-keeping system, regulates various physiological processes through the generation of approximately 24 hour circadian rhythms in gene expression, which are translated into rhythms in metabolism and behavior. It is derived from the Latin roots 'circa' (about) and 'diem' (day) and acts as an important regulator of a wide array of physiological functions including metabolism, sleep, body temperature, blood pressure, endocrine, immune, cardiovascular, and renal function. Consists of two major components: the central clock, residing in the suprachiasmatic nucleus (SCN) of the brain, and the peripheral clocks that are present in nearly every tissue and organ system. Both the central and peripheral clocks can be reset by environmental cues, also known as Zeitgebers (German for 'timegivers'). The predominant Zeitgeber for the central clock is light, which is sensed by retina and signals directly to the SCN. The central clock entrains the peripheral clocks through neuronal and hormonal signals, body temperature and feeding-related cues, aligning all clocks with the external light/dark cycle. Circadian rhythms allow an organism to achieve temporal homeostasis with its environment at the molecular level by regulating gene expression to create a peak of protein expression once every 24 hours to control when a particular physiological process is most active with respect to the solar day. Transcription and translation of core clock components (CLOCK, NPAS2, ARNTL/BMAL1, ARNTL2/BMAL2, PER1, PER2, PER3, CRY1 and CRY2) plays a critical role in rhythm generation, whereas delays imposed by post-translational modifications (PTMs) are important for determining the period (tau) of the rhythms (tau refers to the period of a rhythm and is the length, in time, of one complete cycle). A diurnal rhythm is synchronized with the day/night cycle, while the ultradian and infradian rhythms have a period shorter and longer than 24 hours, respectively. Disruptions in the circadian rhythms contribute to the pathology of cardiovascular diseases, cancer, metabolic syndromes and aging. A transcription/translation feedback loop (TTFL) forms the core of the molecular circadian clock mechanism. Transcription factors, CLOCK or NPAS2 and ARNTL/BMAL1 or ARNTL2/BMAL2, form the positive limb of the feedback loop, act in the form of a heterodimer and activate the transcription of core clock genes and clock-controlled genes (involved in key metabolic processes), harboring E-box elements (5'-CACGTG-3') within their promoters. The core clock genes: PER1/2/3 and CRY1/2 which are transcriptional repressors form the negative limb of the feedback loop and interact with the CLOCK|NPAS2-ARNTL/BMAL1|ARNTL2/BMAL2 heterodimer inhibiting its activity and thereby negatively regulating their own expression. This heterodimer also activates nuclear receptors NR1D1/2 and RORA/B/G, which form a second feedback loop and which activate and repress ARNTL/BMAL1 transcription, respectively. Regulates the circadian expression of ICAM1, VCAM1, CCL2, THPO and MPL and also acts as an enhancer of the transactivation potential of NF-kappaB. Plays an important role in the homeostatic regulation of sleep. The CLOCK-ARNTL/BMAL1 heterodimer regulates the circadian expression of SERPINE1/PAI1, VWF, B3, CCRN4L/NOC, NAMPT, DBP, MYOD1, PPARGC1A, PPARGC1B, SIRT1, GYS2, F7, NGFR, GNRHR, BHLHE40/DEC1, ATF4, MTA1, KLF10 and also genes implicated in glucose and lipid metabolism. Promotes rhythmic chromatin opening, regulating the DNA accessibility of other transcription factors. The CLOCK-ARNTL2/BMAL2 heterodimer activates the transcription of SERPINE1/PAI1 and BHLHE40/DEC1. The preferred binding motif for the CLOCK-ARNTL/BMAL1 heterodimer is 5'-CACGTGA-3', which contains a flanking Ala residue in addition to the canonical 6-nucleotide E-box sequence. CLOCK specifically binds to the half-site 5'-CAC-3', while ARNTL binds to the half-site 5'-GTGA-3'. The CLOCK-ARNTL/BMAL1 heterodimer also recognizes the non-canonical E-box motifs 5'-AACGTGA-3' and 5'-CATGTGA-3'. CLOCK has an intrinsic acetyltransferase activity, which enables circadian chromatin remodeling by acetylating histones and nonhistone proteins, including its own partner ARNTL/BMAL1. Represses glucocorticoid receptor NR3C1/GR-induced transcriptional activity by reducing the association of NR3C1/GR to glucocorticoid response elements (GREs) via the acetylation of multiple lysine residues located in its hinge region. The acetyltransferase activity of CLOCK is as important as its transcription activity in circadian control. Acetylates metabolic enzymes IMPDH2 and NDUFA9 in a circadian manner. Facilitated by BMAL1, rhythmically interacts and acetylates argininosuccinate synthase 1 (ASS1) leading to enzymatic inhibition of ASS1 as well as the circadian oscillation of arginine biosynthesis and subsequent ureagenesis. Drives the circadian rhythm of blood pressure through transcriptional activation of ATP1B1.
-
基因功能參考文獻:
- Clock mRNA expression in the hypoxic-ischemic brain damage (HIBD) group increased at 0 hour after HIBD, decreased at 48 hours after HIBD and increased at 72 hours after HIBD (P<0.05). PMID: 26975828
- A phase advance in rhythmic clock gene expression in the heart and kidney of diabetic rats. PMID: 23864491
- CLOCK protein in the hippocampus but not the suprachiasmatic nucleus plays an important role in long-lasting depressive-like behavior induced by chronic unpredictable stress. PMID: 23263459
- The expressions of Clock and NAT genes in the pineal gland not only show remarkably synchronous endogenous circadian rhythmic changes, but also response to the ambient light signal in a reduced manner. PMID: 15719143
- Clock and Per1 did not differ between behaviorally high and low-responding strains, but there were robust Per2 differences, with high responders having reduced Per2 in ventral tegmental area and substantia nigra PMID: 21775066
- Suprachiamatic clock genes determine the circadian rhythm of prolactin secretion in cervically stimulated rats by regulating dopamine neuronal activity and VIP inputs to the paraventricular nucleus PMID: 19887760
- results indicate a robust circadian expression of clock genes (e.g. Per1 and Bmal1) and the probable existence of a peripheral oscillator in the pancreas. PMID: 15094047
- may regulate intermittent prolactin promoter activity in mammotropes PMID: 15774559
- Data suggest that the different expression patterns observed for Period1, Period nd Clock mRNAs in different brain regions may be attributed to their different physiological roles in these areas. PMID: 15864751
- Circadian expression of the clock gene is demonstrated for the first time in natural killer (NK) cells enriched from rat spleen. PMID: 15944262
- CLOCK showed daily oscillations in different nuclear compartments which differed significantly between adult & old rats; this unusual distribution during aging could be related to the prolonged diurnal activity of old rats or to altered nuclear pathways PMID: 16208722
- clock gene expression within dopaminergic neurons may act to modulate diurnal rhythms of dopamine release PMID: 16373438
- test the importance of Clock variants in explaining diversity among rat strains in the expression of phenotypes, such as blood pressure, subject to circadian oscillation PMID: 16376516
- rhythms in clock gene expression for the most part develop postnatally PMID: 16603678
- We suggest that the circadian Clock protein may help coordinate the sensing of energy expenditure with energy supply. PMID: 17097616
- These results show that interferon-gamma can alter the electrical properties and circadian clock gene expression in suprachiasmatic nucleus neurons. PMID: 18375864
- rhythmic melatonin administration has an effect on clock gene expression in the suprachiasmatic nucleus and the heart of hypertensive TGR(mRen2)27 rats PMID: 19633447
顯示更多
收起更多
-
亞細胞定位:Cytoplasm. Nucleus. Cytoplasm, cytosol.
-
組織特異性:Expressed in the suprachiasmatic nucleus (SCN), and in the piriform cortex (PC).
-
數據庫鏈接:
Most popular with customers
-
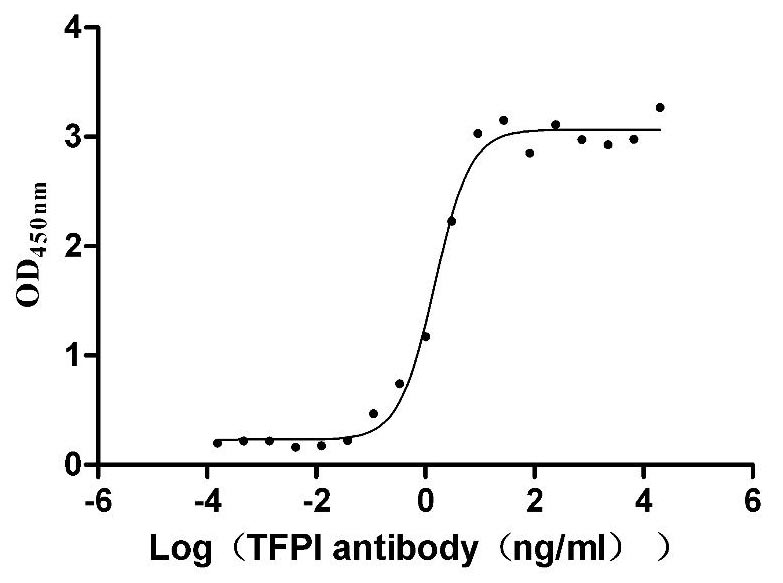
Recombinant Human Tissue factor pathway inhibitor (TFPI), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Intestinal-type alkaline phosphatase (ALPI) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
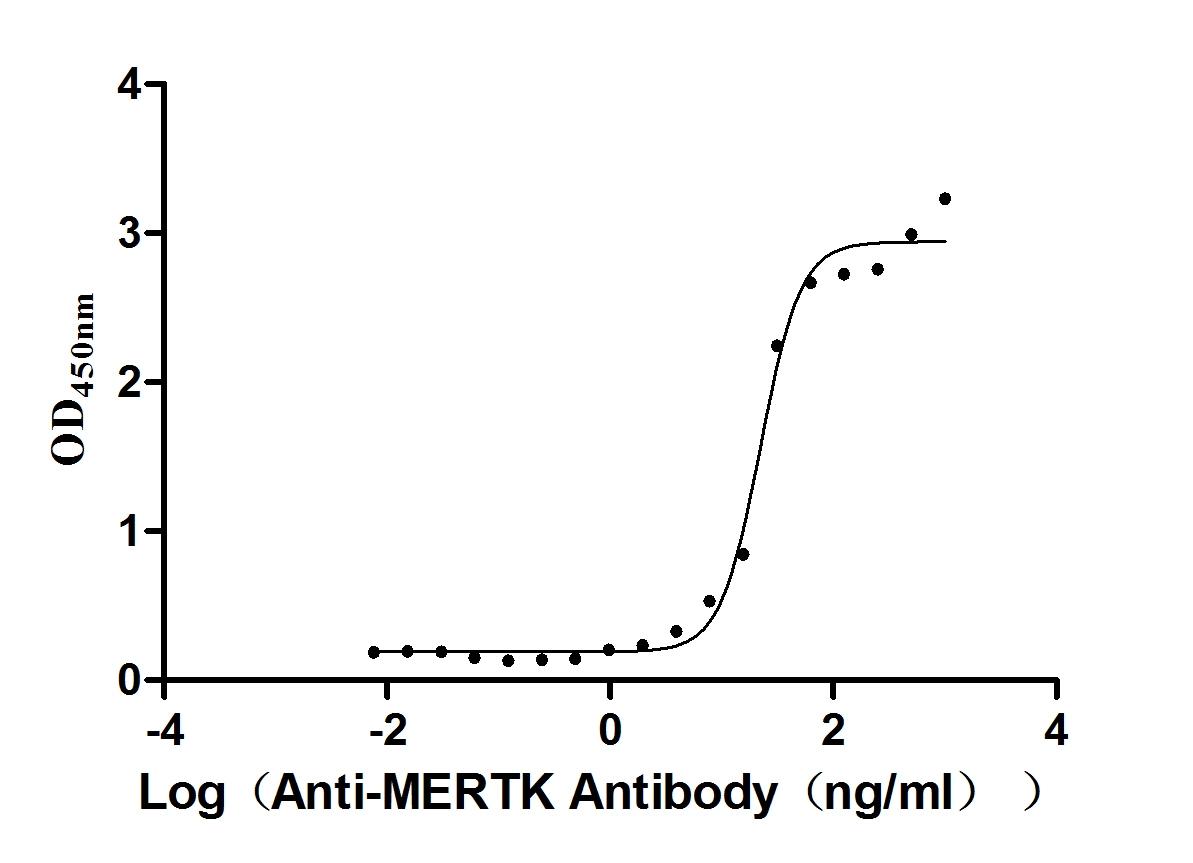
Recombinant Mouse Tyrosine-protein kinase Mer (Mertk), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
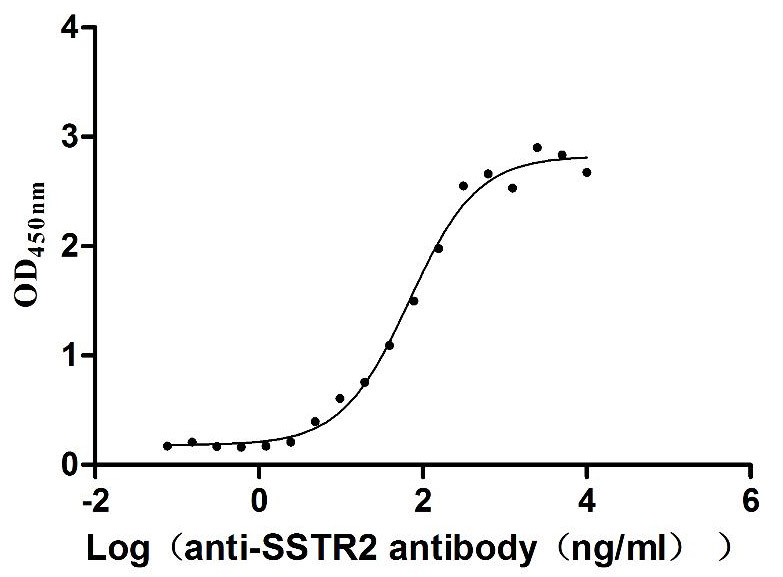
Recombinant Human Somatostatin receptor type 2 (SSTR2)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
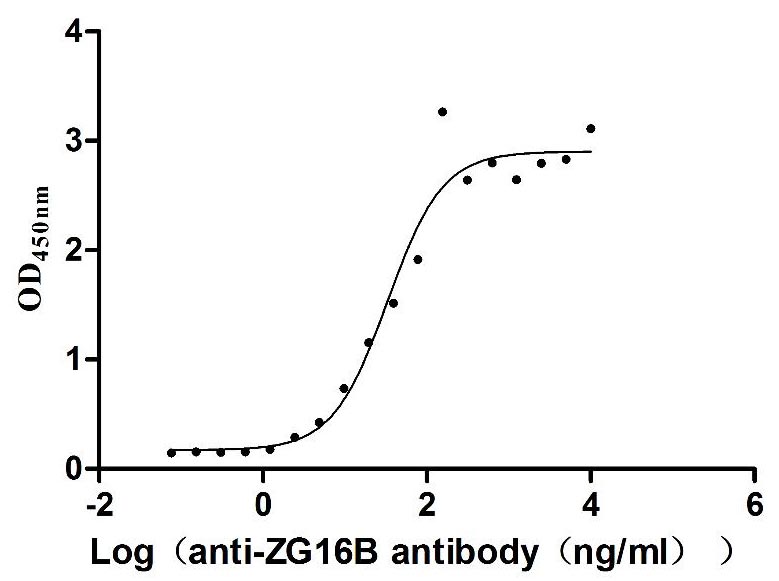
Recombinant Human Pancreatic adenocarcinoma up-regulated factor (ZG16B) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
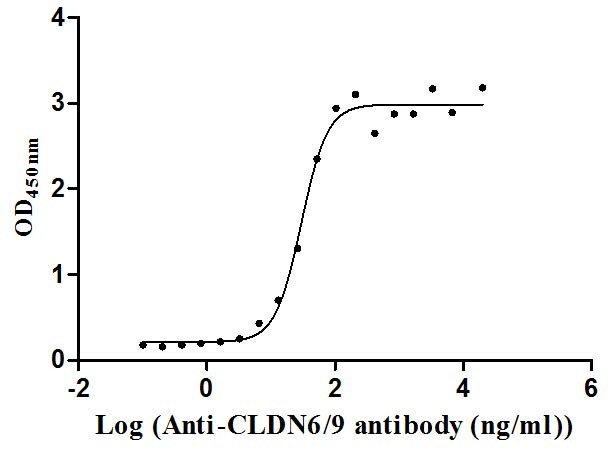
Recombinant Human Claudin-9 (CLDN9)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Mouse Complement component C1q receptor (Cd93), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
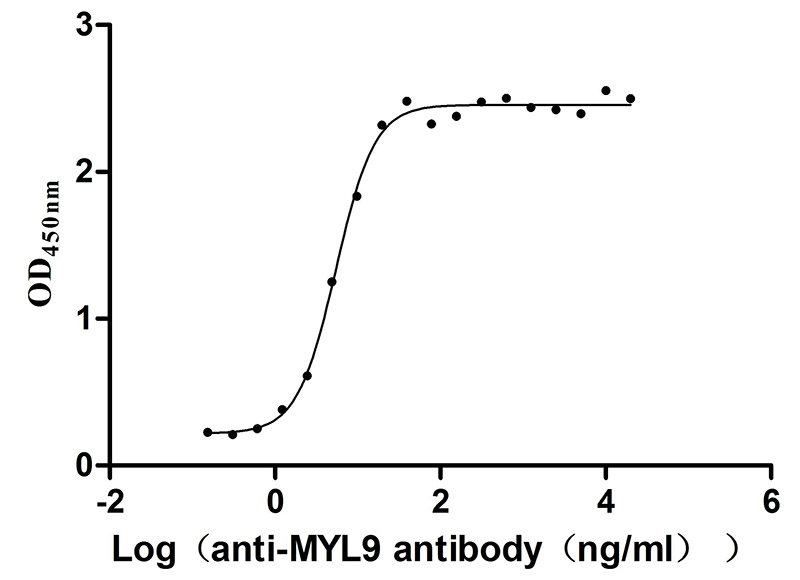
Recombinant Human Myosin regulatory light polypeptide 9 (MYL9) (Active)
Express system: Yeast
Species: Homo sapiens (Human)









