Recombinant Mouse Microprocessor complex subunit DGCR8 (Dgcr8)
-
中文名稱:小鼠Dgcr8重組蛋白
-
貨號:CSB-EP866818MO-B
-
說明書:
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:Dgcr8
-
Uniprot No.:
-
別名:Dgcr8Microprocessor complex subunit DGCR8; DiGeorge syndrome critical region 8 homolog; Gy1
-
種屬:Mus musculus (Mouse)
-
蛋白長度:full length protein
-
表達區域:1-773
-
氨基酸序列METYESPSPL PREPAGEAMM ENRACPFQVL PHEQSPPPPL QTSSDAEVMD VGSGGDGQSE PPADDPFNFY GASLLSKGSF SKGRLLIDPN CSGHSPRTAR HAPAVRKFSP DLKLLKDVKI SVSFTESCRS KDRKVLYTGV ERSTRPECGQ LLSPVSGDVH ACPFGGSVGN GVGLGGESAD KKDEENELDQ EKRVEYAVLD ELEDFTDNLE LDEEGTGGFT AKAIVQRDRV DEEALNFSYE DDFDNDVDAL LEEGLCAPKK RRMEEKYGGD SDHPSDGETS VQPMMTKIKT VLKSRGRPPT EPLPDGWIMT FHNSGVPVYL HRESRVVTWS RPYFLGTGSI RKHDPPLSSI PCLHYKKMKD NEEREQNCDL APSGEVSPVK PLGRSAELDF PLEEPDSMGG DSGSMDEKDP LGAEAAAGAL GQVKAKVEVC KDESVDLEEF RNYLEKRFDF EQVTVKKFRT WAERRQFNRE MKRKQAESER PILPANQKLI TLSVQDAPTK KEFVINPNGK SEVCILHEYM QRVLKVRPVY NFFECENPSE PFGASVTIDG VTYGSGTASS KKLAKNKAAR ATLEILIPDF VKQTSEEKPK DSEELEYFNH ISIEDSRVYE LTSKAGLLSP YQILHECLKR NHGMGDTSIK FEVVPGKNQK SEYVMACGKH TVRGWCKNKR VGKQLASQKI LQLLHPHVKN WGSLLRMYGR ESSKMVKQET SDKSVIELQQ YAKKNRPNLH ILSKLQEEMK RLAAEREETR KKPKMSIVAS AQPGGEPLCT VDV
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Component of the microprocessor complex that acts as a RNA- and heme-binding protein that is involved in the initial step of microRNA (miRNA) biogenesis. Component of the microprocessor complex that is required to process primary miRNA transcripts (pri-miRNAs) to release precursor miRNA (pre-miRNA) in the nucleus. Within the microprocessor complex, DGCR8 function as a molecular anchor necessary for the recognition of pri-miRNA at dsRNA-ssRNA junction and directs DROSHA to cleave 11 bp away form the junction to release hairpin-shaped pre-miRNAs that are subsequently cut by the cytoplasmic DICER to generate mature miRNAs. The heme-bound DGCR8 dimer binds pri-miRNAs as a cooperative trimer (of dimers) and is active in triggering pri-miRNA cleavage, whereas the heme-free DGCR8 monomer binds pri-miRNAs as a dimer and is much less active. Both double-stranded and single-stranded regions of a pri-miRNA are required for its binding. Specifically recognizes and binds N6-methyladenosine (m6A)-containing pri-miRNAs, a modification required for pri-miRNAs processing. Involved in the silencing of embryonic stem cell self-renewal.
-
基因功能參考文獻:
- The Ddx6 knockout cells were phenotypically and molecularly similar to cells lacking all microRNAs (Dgcr8 knockout ESCs). These data show that the loss of DDX6 can separate the two canonical functions of microRNAs: translational repression and transcript destabilization PMID: 30044225
- Deletion of DGCR8 results in loss of vascular reactivity, reduced blood pressure and neointima formation. PMID: 29362439
- these results strongly indicate that DGCR8-dependent generation of miR-22 is essential for bone formation PMID: 28739418
- We show that Dgcr8 mutations induce an earlier and stronger phenotype in the developing nervous system compared to Dicer mutants and that miRNA-independent functions of DGCR8 are critical for corticogenesis. PMID: 28232627
- this study shows that B-cell specific cre-mediated DGCR8 deletion blocks B-cell development at the transition from the pro-B to the pre-B cell stage PMID: 27641147
- DGCR8 in the modulates of the alternative splicing of Tcf7l1 mRNA in addition to its established function in microRNA biogenesis, controlling embryonic stem cell exit from pluripotency. PMID: 28100686
- we suggest that DGCR8-dependent canonical microRNAs are essential for uterine development and physiological processes such as proper immune modulation, reproductive cycle, and steroid hormone responsiveness in mice. PMID: 26833131
- Deletion of the Dgcr8 gene inhibits tumor progression in a Pten-knockout mouse model of prostate cancer PMID: 26206718
- DGCR8 is required for the progression, but not initiation, of Akt induced prostate cancer in vivo. PMID: 26206718
- DGCR8 is required for microRNA biogenesis and normal mouse embryonic stem cell proliferation and differentiation. PMID: 17259983
- These results reveal a new pathway in the DNA damage response wherein ABL-dependent tyrosine phosphorylation of DGCR8 stimulates the processing of selective primary miRNAs. PMID: 26126715
- Conditional gene deletion of the essential miRNA-processing enzyme Dgcr8 in the developing renal tubular system results in severe developmental defects and kidney failure. PMID: 25881298
- Dgcr8 is responsible for modulation of gene expression programs underlying myelin formation and maintenance as well as suppression of an injury-related gene expression program PMID: 26272614
- Dgcr8 mutant mice, which have a defective miRNA pathway while retaining an intact endo-siRNA pathway, were infertile and displayed cumulative defects in meiotic and haploid phases of spermatogenesis, resulting in oligo-, terato-, and azoospermia. PMID: 25244517
- Adipose tissue-specific Dgcr8 knockout mice displayed enlarged but pale interscapular brown fat with decreased expression of genes characteristic of brown fat and were intolerant to cold exposure. PMID: 25008181
- Data indicate that DiGeorge syndrome critical region gene 8 (DGCR8)-dependent miRs are indispensable for osteoclastic control of bone metabolism. PMID: 24420069
- This suggests that Dgcr8-microRNA-Drd2-dependent thalamocortical disruption is a pathogenic event underlying schizophrenia-associated psychosis. PMID: 24904170
- These data reveal a role for DeltaNp63 in the transcriptional regulation of DGCR8 to reprogram adult somatic cells into multipotent stem cells. PMID: 24449888
- Data suggest that Dgcr8-mediated regulation of microRNA is likely to underlie Cxcr4/Cxcl12 signaling and associated neurodevelopmental defects. PMID: 24101523
- The results of this study suggested that robust alterations in synaptic properties within the prefrontal cortex, which to a large extent but not entirely can be accounted for by Dgcr8 deficiency. PMID: 24027283
- Several schizophrenia-associated genes were downregulated in the hippocampus of Dgcr8(+/-) mice, and one of them, insulin-like growth factor 2 PMID: 23719809
- a novel candidate for Dgcr8-independent microRNA genesis PMID: 22912678
- Deletion of Dgcr8 results in a severe deficit in inhibitory synaptic transmission and reduction of inhibitory synapses in pyramidal neurons . PMID: 22728723
- DGCR8 gene is required for vascular development through the regulation of VSMC proliferation, apoptosis, and differentiation. PMID: 22511778
- Dgcr8 controls neural crest cells survival in cardiovascular development. PMID: 22138056
- loss of Dicer1 resulted in an earlier lethality, more severe structural abnormalities, and increased apoptosis relative to that from Dgcr8 loss PMID: 21712401
- In this study, we show that Dgcr8+/- mice display reduced expression of a subset of microRNAs in the prefrontal cortex PMID: 21466685
- The effect of Dgcr8 deficiency was determined on the structure and function of cortical circuits by assessing their laminar organization. PMID: 21368174
- ING1 binds to the DGCR8 promoter and controls its transcription through chromatin regulation. ING1 and DGCR8 can cooperate in restraining proliferation. PMID: 20179197
- a Microprocessor, containing the RNA binding protein Dgcr8 and RNase III enzyme Drosha, is responsible for processing primary microRNAs to precursor microRNAs PMID: 19759829
- Analysis of mouse knockout ES cells shows that DGCR8 is essential for biogenesis of miRNAs PMID: 17259983
- Haploinsufficiency of the Dgcr8 gene is associated with behavioral and neuronal deficits PMID: 18469815
- most abundantly expressed skin microRNAs are dependent on both Dicer and DGCR8 PMID: 19114655
- Cardiomyocyte-specific deletion of dgcr8, a gene required for microRNA biogenesis, revealed a fully penetrant phenotype that begins with left ventricular malfunction progressing to a dilated cardiomyopathy and premature lethality PMID: 19679836
顯示更多
收起更多
-
亞細胞定位:Nucleus. Nucleus, nucleolus.
-
組織特異性:Ubiquitously expressed.
-
數據庫鏈接:
Most popular with customers
-
-AC1.jpg)
Recombinant Human Claudin-18.2 (CLDN18.2)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
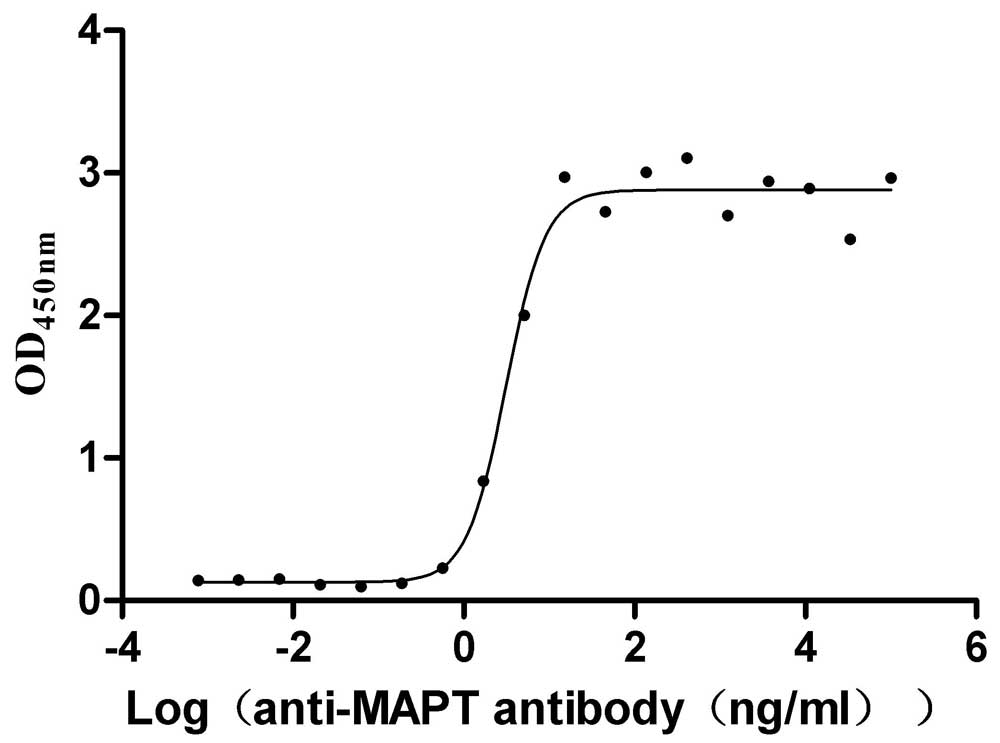
Recombinant Macaca mulatta Microtubule-associated protein tau (MAPT) (Active)
Express system: Mammalian cell
Species: Macaca mulatta (Rhesus macaque)
-
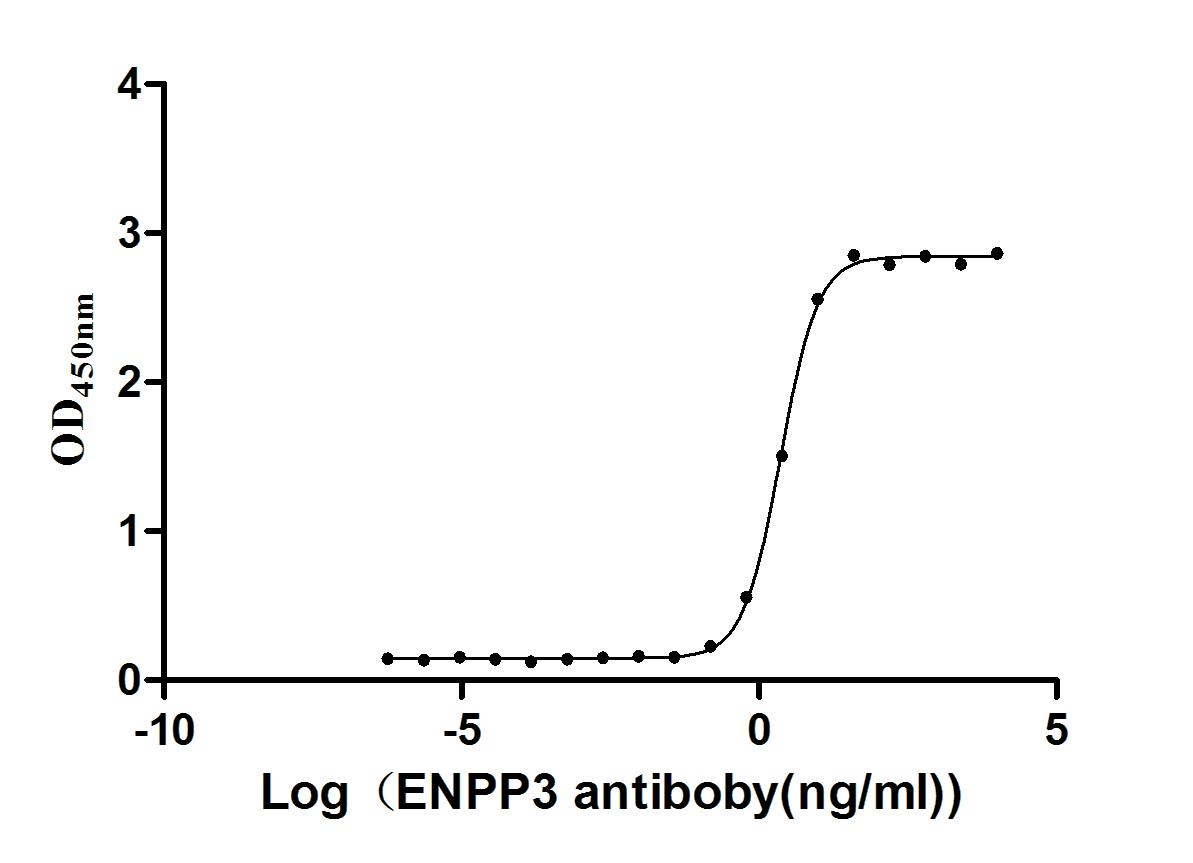
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
f4-AC1.jpg)
Recombinant Human Claudin-6 (CLDN6)-VLPs, Fluorescent (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









