Recombinant Mouse Lymphocyte antigen 96 (Ly96)
In Stock-
中文名稱:小鼠Ly96重組蛋白
-
貨號:CSB-EP013254MO
-
規格:¥1836
-
圖片:
-
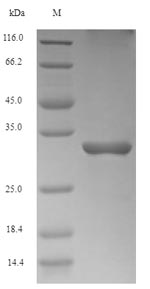 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產品詳情
-
純度:Greater than 90% as determined by SDS-PAGE.
-
基因名:
-
Uniprot No.:
-
別名:Ly96; Esop1; Md2; Lymphocyte antigen 96; Ly-96; ESOP-1; Protein MD-2
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length of Mature Protein
-
來源:E.coli
-
分子量:32.4kDa
-
表達區域:19-160aa
-
氨基酸序列EKQQWFCNSSDAIISYSYCDHLKFPISISSEPCIRLRGTNGFVHVEFIPRGNLKYLYFNLFISVNSIELPKRKEVLCHGHDDDYSFCRALKGETVNTSIPFSFEGILFPKGHYRCVAEAIAGDTEEKLFCLNFTIIHRRDVN
Note: The complete sequence may include tag sequence, target protein sequence, linker sequence and extra sequence that is translated with the protein sequence for the purpose(s) of secretion, stability, solubility, etc.
If the exact amino acid sequence of this recombinant protein is critical to your application, please explicitly request the full and complete sequence of this protein before ordering. -
蛋白標簽:N-terminal 6xHis-SUMO-tagged
-
產品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:If the delivery form is liquid, the default storage buffer is Tris/PBS-based buffer, 5%-50% glycerol.
Note: If you have any special requirement for the glycerol content, please remark when you place the order.
If the delivery form is lyophilized powder, the buffer before lyophilization is Tris/PBS-based buffer, 6% Trehalose. -
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:3-7 business days
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet & COA:Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Binds bacterial lipopolysaccharide (LPS). Cooperates with TLR4 in the innate immune response to bacterial lipopolysaccharide (LPS), and with TLR2 in the response to cell wall components from Gram-positive and Gram-negative bacteria. Enhances TLR4-dependent activation of NF-kappa-B. Cells expressing both LY96 and TLR4, but not TLR4 alone, respond to LPS.
-
基因功能參考文獻:
- We conclude that MD2 is a significant contributor in the Ang II-induced kidney inflammatory injury in chronic renal diseases. PMID: 28322341
- Blockade of MD2 prevents obesity-induced inflammation and nephropathy. PMID: 28767204
- MD2 is essential to obesity-related cardiac hypertrophy through activating JNK/ERK and NF-kappaB-dependent cardiac inflammatory pathways. Targeting MD2 would be a therapeutic approach to prevent obesity-induced cardiac injury and remodeling. PMID: 28965884
- RTFs contribute to the regulation of LPS-induced inflammatory response in RAW264.7 cells through TLR4/MD-2 mediated NF-kappaB and JNK pathway. It PMID: 27235587
- This study provides evidence that MD2 plays a key role in the pathogenesis of retinal I/R damage. PMID: 29111459
- Data suggest that C4bp prevents interaction between Tlr4/MD-2 and its ligand; C4bp does not appear to interact with Tlr3; C4bp binds to macrophage surface Tlr4 and inhibits Tlr/Tlr ligand interaction, thereby inhibiting Tlr4 activation. (C4bp = complement component 4 binding protein; Tlr = toll-like receptor; MD-2 = myeloid differentiation protein-2) PMID: 28542817
- MD2 plays an important role in induction of allergic sensitization to cat dander and common pollens relevant to human allergic diseases. PMID: 26586036
- Oxidative stress in retinal ischemia-reperfusion injury activates TLR4 signaling via MD2. PMID: 28063877
- Neoseptin-3 and lipid A form dissimilar molecular contacts to achieve receptor activation; hence strong TLR4/MD-2 agonists need not mimic LPS PMID: 26831104
- Here we demonstrate that cholesterol binds to myeloid differentiation-2 (MD-2), a TLR4 ancillary molecule. PMID: 26806306
- MD-2 is a critical regulator of the establishment of allergic airway sensitization to HDM in mice. Serum MD-2 may represent a potential biomarker for the amplification of allergic sensitization and allergic inflammation. PMID: 26344079
- Data show that myeloid differentiation factor 2 (MD-2) binds specifically to disulfide isoform of box protein 1, high mobility group (HMGB1) to facilitate toll-like receptor 4 (TLR4)-dependent signaling. PMID: 25559892
- Carbon monoxide treatment reduces the expression of the TLR4/MD2 complex on the surface of myeloid cells, which renders them resistant to lipopolysaccharide priming in vitro, as well as in vivo in a model of endotoxic shock. PMID: 25179131
- Mechanistically, engagement of MD-2 by PTX3-opsonized Aspergillus conidia activated the TLR4/Toll/IL-1R domain-containing adapter inducing IFN-beta-dependent signaling pathway converging on IL-10. PMID: 25049357
- SAA3 directly binds MD-2 and activates the MyD88-dependent TLR4/MD-2 pathway. PMID: 23858030
- Monophosphoryl lipid A is unable to efficiently form TLR4/MD-2 heterotetramers, but it still needs heterotetramer formation for the full extent of signaling it is able to achieve. PMID: 23638128
- Data show that rifampin binds to myeloid differentiation protein 2 (MD-2), the key coreceptor for innate immune TLR4. PMID: 23568774
- Gb4 is an endogenous ligand for TLR4-MD-2 and is capable of attenuating LPS toxicity, indicating the possibility for its therapeutic application in endotoxin shock. PMID: 23471986
- Data provide structural evidence of the agonistic property of lipid IVa on TLR4/MD-2 and deepen understanding of the ligand binding and dimerization mechanism by the structurally diverse Lipopolysaccharide (LPS) variants. PMID: 22532668
- GL and ILG modulate the TLR4/MD-2 complex at the receptor level, leading to suppress LPS-induced activation of signaling cascades and cytokine production PMID: 22422925
- Data show that morphine binds to an accessory protein of Toll-like receptor 4 (TLR4), myeloid differentiation protein 2 (MD-2), thereby inducing TLR4 oligomerization and triggering proinflammation. PMID: 22474354
- Data show that LPS priming of tolerant FLDCs inhibited the up-regulation of TLR4/MD-2 expression. PMID: 21802073
- According to our murine TLR4/MD-2-activation model, the two phosphates on lipid A were predicted to interact extensively with the two positively charged patches on mouse TLR4activation PMID: 21865549
- Demonstrate a novel, critical role for MD-2 and TLR4 through NADPH activation in liver steatosis, and fibrosis in a NASH model in mice. PMID: 21233280
- species-specific activation of lipid IV(A) PMID: 20592019
- Neutralizing toll-like receptor 4/myeloid differentiation protein-2 is highly efficacious in protecting against bacterial infection-induced toxemia. PMID: 17947685
- MD-2-mediated ionic interactions between lipid A and TLR4 are essential for receptor activation PMID: 20018893
- MD-2 in complex with toll-like receptor 4 mediates signal transduction induced by the amino acid-containing bacterial lipid, flavolipin. PMID: 11884465
- MD-2 is essential for correct intracellular distribution and LPS-recognition of TLR4. PMID: 12055629
- By alanine-scanning mutagenesis of MD-2, important amino acid residues have been identified that confer lipopolysaccharide and taxol responsiveness on TLR4 and enable formation of cell surface TLR4-MD-2 complex recognized by specific monoclonal antibody. PMID: 12496426
- TLR4 and its partner molecule MD-2 may play an important role in Kupffer cell activation and ischemia-reperfusion injury. PMID: 15334694
- Results show that the N-terminal region of toll-like receptor 4 is essential for association with MD-2, which is required for the cell surface expression and hence the responsiveness to lipopolysaccharide. PMID: 15337750
- Collectively, MD-2 is essential for the recognition of LPS by TLR4 but not for that of PGN by TLR2 of mast cells. PMID: 15369778
- Results indicate that amino acid residues 57, 61, and 122 of mouse MD-2 are critical to determine the agonist-antagonist activity of lipid IVa and suggest that these amino acid residues may be involved in the discrimination of lipid A structure. PMID: 16407172
- agonistic mAb to Toll-Like Receptor 4 (TLR4)/MD-2 protected mice from lipopolysaccharide/d-galactosamine-induced acute lethal hepatitis by delivering a protective signal activating NF-kappaB through TLR4/MD-2 PMID: 16547261
- This study shows regulatory roles for MD-2 in initiating and terminating ligand-induced TLR4 oligomerization. PMID: 16670331
- a unique complex of TLR4, MD-2, and CD44 recognizes hyaluronan in signaling tissue injury PMID: 17400552
- Data show that lipopolysaccharides act through both MyD88-dependent and -independent TLR4/MD2 signaling pathways to directly inhibit GHR gene expression. PMID: 17601656
- Based on structural analysis and mutagenesis experiments on MD-2 and TLR4, we propose a model of TLR4-MD-2 dimerization induced by LPS. PMID: 17803912
- The lower expression of the CD14 and TLR-4/MD-2 receptors may be partly responsible for the immunodeficiency observed in the malnourished mice. PMID: 17950615
- MD-2 is a newly recognized type II acute-phase reactant, an opsonin for Gram-negative bacteria, and a cofactor essential for the activation of TLR4-expressing cells. PMID: 18056837
- role of the TLR4/MD-2 signaling axis in bacterial recognition by phagocytes PMID: 18203953
- Data compare the inflammatory potency of two types of Neisseria meningitidis endotoxins in lungs: wild type (hexaacylated, LOS(wt)) and mutant type (pentaacylated, LOS(msbB)), and describe the importance of MD-2 in endotoxin responses in lungs in vivo. PMID: 18203970
- The species-specific difference between human and murine MD-2 activation of TLR4 by PTX can be explained by alterations of surface charge distribution (i.e. electrostatic potential), binding pocket size, and the locus of PTX binding within the MD-2 pocket PMID: 18650420
- Myeloid differentiation protein-2 has an important role in the CD14-independent LPS-mediated cascade of neutrophil influx PMID: 18988922
- the leucine at position 815 is required for the normal maturation of TLR4 and for formation of the TLR4.MD-2 complex. PMID: 19064998
- These findings reveal novel roles of lysines 122, 125, and 58 in human MD-2 that contribute to the functional differences between human and murine MD-2 and, potentially, to differences in the sensitivity of humans and mice to endotoxin. PMID: 19783674
顯示更多
收起更多
-
亞細胞定位:Secreted, extracellular space. Secreted.
-
組織特異性:Highly expressed in spleen, bone marrow, thymus, liver, kidney, ovary and decidua. Detected at lower levels in testis, small intestine and skin.
-
數據庫鏈接:
Most popular with customers
-
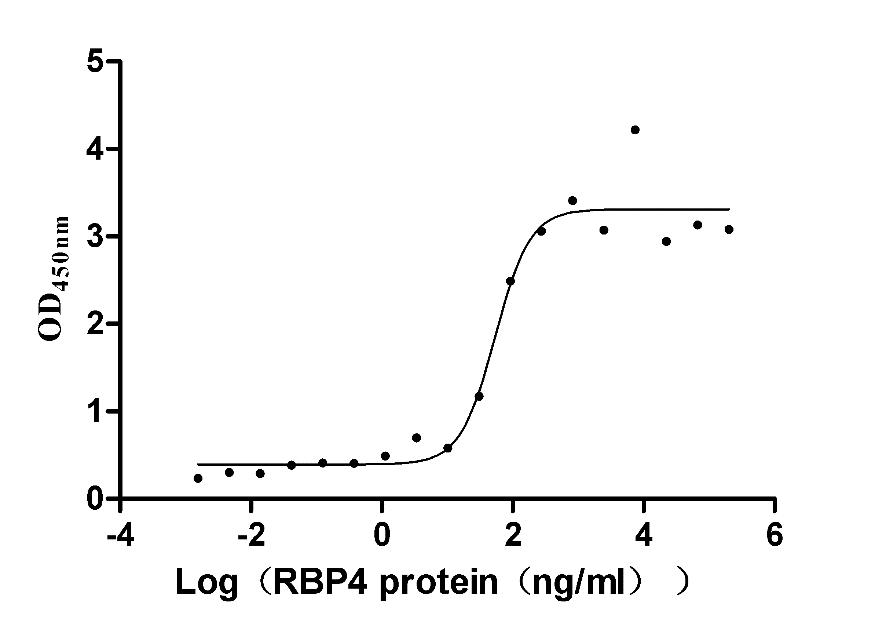
Recombinant Mouse Retinol-binding protein 4 (Rbp4) (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
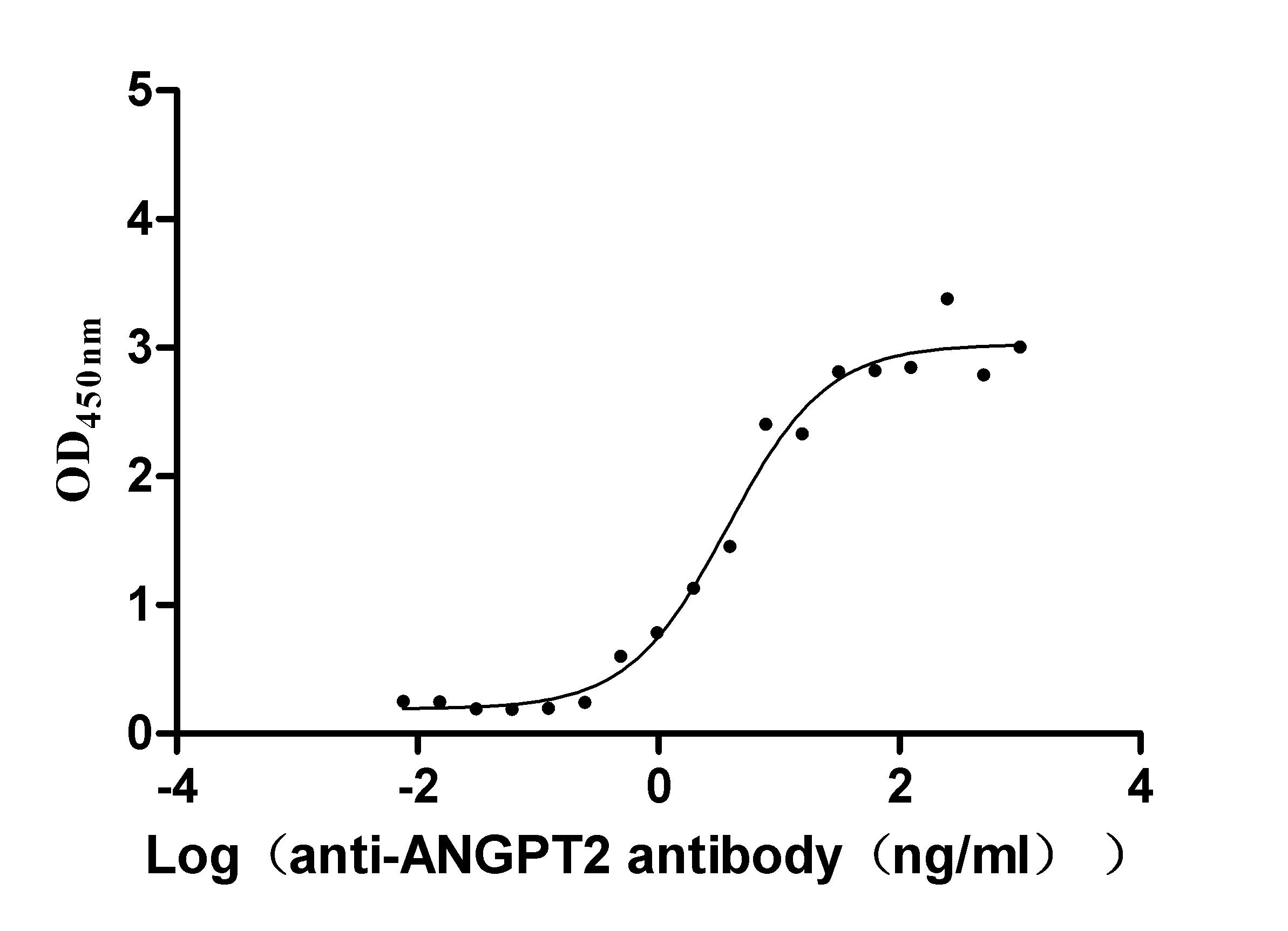
Recombinant Dog Angiopoietin-2 (ANGPT2) (Active)
Express system: Mammalian cell
Species: Canis lupus familiaris (Dog) (Canis familiaris)
-
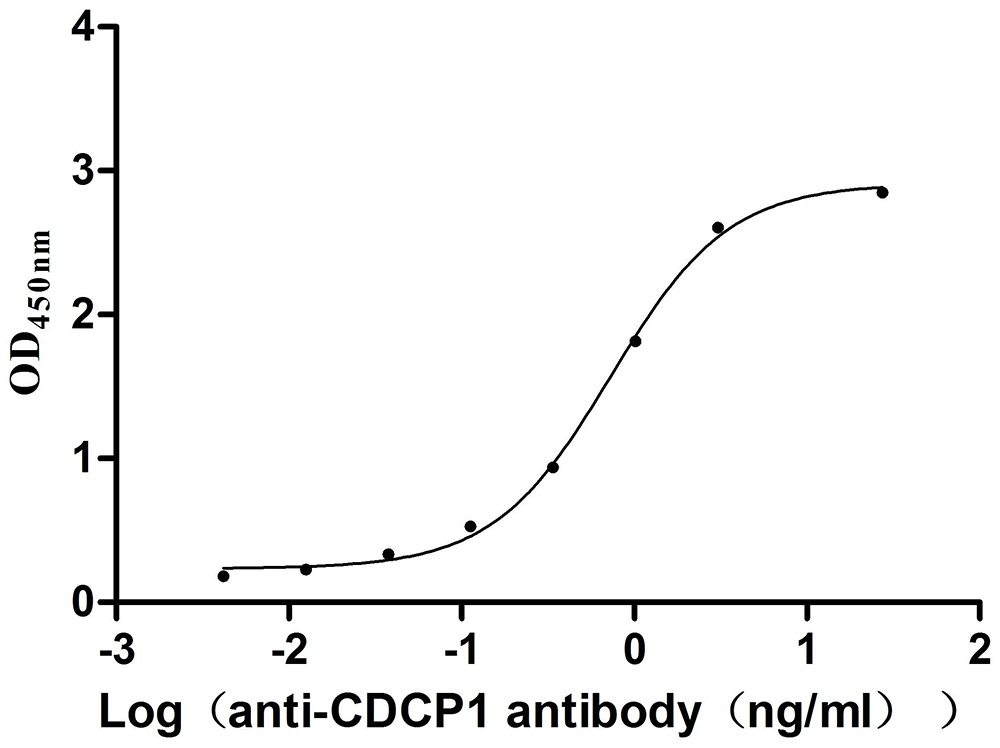
Recombinant Mouse CUB domain-containing protein 1 (Cdcp1), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
Recombinant Human Carcinoembryonic antigen-related cell adhesion molecule 8(CEACAM8) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Cadherin-1(CDH1),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









