Recombinant Arabidopsis thaliana E3 ubiquitin-protein ligase COP1 (COP1), partial
-
中文名稱:Recombinant Arabidopsis thaliana E3 ubiquitin-protein ligase COP1(COP1) ,partial
-
貨號:CSB-YP337358DOA
-
說明書:
-
規(guī)格:
-
來源:Yeast
-
其他:
-
中文名稱:Recombinant Arabidopsis thaliana E3 ubiquitin-protein ligase COP1(COP1) ,partial
-
貨號:CSB-EP337358DOA
-
說明書:
-
規(guī)格:
-
來源:E.coli
-
其他:
-
中文名稱:Recombinant Arabidopsis thaliana E3 ubiquitin-protein ligase COP1(COP1) ,partial
-
貨號:CSB-EP337358DOA-B
-
說明書:
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Arabidopsis thaliana E3 ubiquitin-protein ligase COP1(COP1) ,partial
-
貨號:CSB-BP337358DOA
-
說明書:
-
規(guī)格:
-
來源:Baculovirus
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:COP1
-
Uniprot No.:
-
別名:COP1; At2g32950; T21L14.11; E3 ubiquitin-protein ligase COP1; EC 2.3.2.27; Constitutive photomorphogenesis protein 1; RING-type E3 ubiquitin transferase COP1
-
種屬:Arabidopsis thaliana (Mouse-ear cress)
-
蛋白長度:Partial
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder Warning: in_array() expects parameter 2 to be array, null given in /www/web/cusabio_cn/public_html/caches/caches_template/default/content/show_product_protein.php on line 662
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評價

靶點詳情
-
功能:E3 ubiquitin-protein ligase that acts as a repressor of photomorphogenesis and as an activator of etiolation in darkness. E3 ubiquitin ligases accept ubiquitin from an E2 ubiquitin-conjugating enzyme in the form of a thioester and then directly transfers the ubiquitin to targeted substrates. Represses photomorphogenesis in darkness by mediating ubiquitination and subsequent proteasomal degradation of light-induced transcription factors such as HY5, HYH and LAF1. Down-regulates MYB21, probably via ubiquitination process. Light stimuli abrogate the repression of photomorphogenesis, possibly due to its localization to the cytoplasm. Could play a role in switching between skotomorphogenetic and photomorphogenetic pathways. Mediates the ubiquitination-dependent degradation of HY5 in the darkness during seedling development (e.g. hypocotyl growth). Represses CIP7 in darkness.
-
基因功能參考文獻:
- COL12 is a substrate of the COP1. PMID: 29187570
- plays a prominent role in drought stress tolerance PMID: 30139551
- Mutation of the coiled-coil domain in COP1, which prevents dimer formation, impairs COP1 function in coordinating flowering time. PMID: 29273730
- Nitrate Reductases levels are negatively regulated by COP1 and ammonium. PMID: 29662028
- Data indicate that light is directly linked with the inducer of CBF Expression (ICE) transcription factors (ICE)-directed signaling module, via the CONSTITUTIVE PHOTOMORPHOGENIC1 (COP1)-mediated protein surveillance system, in the modulation of stomatal development. PMID: 29070509
- COP1/SPA complex associates with and stabilizes PHYTOCHROME INTERACTING FACTOR 3 (PIF3) to repress photomorphogenesis in the dark. PMID: 28292892
- High COP1 expression is associated with defense against turnip crinkle virus. PMID: 29513740
- COP1 mediates dark-specific degradation of microtubule-associated protein WDL3 in regulating Arabidopsis hypocotyl elongation. PMID: 29087315
- Thermal activation of COP1 enables coincidence between warm temperature signaling and circadian rhythms, which allows plants to gate hypocotyl thermomorphogenesis at the most profitable time at warm temperatures. PMID: 28418582
- ABI4 and HY5 antagonistically regulate the expression of CONSTITUTIVE PHOTOMORPHOGENIC1 (COP1) and the subsequent greening process. In turn, ABI4 and HY5 are targeted for degradation by COP1 in the light and dark, respectively, to ensure a proper interplay of ABI4 and HY5 actions during seedling de-etiolation. PMID: 27255835
- Data indicate localization of UVR8 signaling in the nucleus and a dual role for COP1 in the regulation of UV-B-induced UVR8 nuclear accumulation and in UVR8-mediated UV-B signaling. PMID: 27407149
- SPA proteins have a dual role: (1) they are required for light-responsiveness of COP1 subcellular localization, and (2) they promote COP1 activity in darkness in a fashion that is independent of the nuclear import/nuclear retention of COP1. PMID: 28536102
- The alpha2-cop mutant displayed defects in plant growth, including small rosettes, stems and roots and mislocalization of p24delta5, a protein of the p24 family containing a C-terminal dilysine motif involved in COPI binding. The alpha2-cop mutant also exhibited abnormal morphology of the Golgi apparatus. PMID: 28025315
- Exposure to blue light is required for an in vivo-association of CRY1 and CRY2 with COP1. PMID: 28991901
- DHU1 negatively regulates UV-B signaling via its direct interaction with COP1 and RUP1 (At5g52250). PMID: 28735869
- BBX21 is a pivotal component involved in the COP1-HY5 regulatory hub. PMID: 27325768
- Meanwhile, transcript levels of ABA biosynthesis genes are higher in cip1-1 than in the wild-type. These results suggested that CIP1 is positively involved in ABA response. PMID: 27372427
- salt stress and ethylene antagonistically regulate nucleocytoplasmic partitioning of COP1. PMID: 26850275
- Our genetic and biochemical studies identify a function for SIZ1 in photomorphogenesis and reveal a novel SUMO-regulated ubiquitin ligase, COP1, in plants. PMID: 27128446
- Data indicate a coordinated regulation of Arabidopsis proteins SHW1, COP1, and HY5 in seedling development. PMID: 26474641
- The COP1 role in CONSTANS protein degradation during photoperiodic flowering PMID: 26358558
- CSU2 interacts with COP1 via the coiled-coil domain association. CSU2 negatively regulates COP1 E3 ubiquitin ligase activity. [COP1] PMID: 26714275
- propose that light perceived by phytochromes causes a switch in the ubiquitination activity of COP1/SPA2 from ubiquitinating downstream substrates to ubiquitinating SPA2, which subsequently causes a repression of COP1/SPA2 function PMID: 26368289
- CUL4(COP1-SPA) E3 ubiquitin ligase is necessary for the light-induced degradation of PIF1 in Arabidopsis. PMID: 26037329
- genetic analyses with transgenes expressing a genomic pmARI12:ARI12-GFP construct confirm the epistatic interaction between COP1 and ARI12 in growth responses to high fluence rate UV-B. PMID: 25817546
- AN3 may act with other proteins that bind to COP1 promoter to promote anthocyanin accumulation and inhibit light-induced root elongation. PMID: 25256341
- Two distinct domains of the UVR8 photoreceptor interact with COP1 to initiate UV-B signaling in Arabidopsis PMID: 25627067
- cytoplasmic partitioning of COP1 under light is essential to protect HYL1 against protease X. PMID: 25532508
- It was shown show that mutations in the key repressor of light signaling, the COP1/SPA complex, cause a strong hyperaccumulation of anthocyanins not only under normal light but also under high light conditions. PMID: 25482806
- Light and COP1 regulate level of overexpressed DET1 protein PMID: 25575996
- Analysis of BBX22 degradation kinetics shows that it has a short half-life under both dark and light conditions. COP1 mediates the BBX22 degradation in the dark. Although dispensable in the dark, HY5 contributes to the BBX22 degradation in the light. PMID: 21427283
- COP1 is revealed as a potential coordinator of cytoskeletal and electrophysiological activities required for guard cell function PMID: 25151660
- Based on our molecular and biochemical evidence, we propose that the UVR8-COP1 affinity in plants critically determines the photomorphogenic UV-B signal transduction coupling with UVR8-mediated UV-B light perception. PMID: 24651064
- COP1 inactivation involved in rapid light-induced responses were compared to that of nuclear HY5. PMID: 24434030
- our investigation demonstrates that the COP1-HY5 complex is a novel integrator that plays an essential role in ethylene-promoted hypocotyl growth in the light. PMID: 24348273
- The MID protein plays a role in COP1/SPA1-controlled repression of flowering under short-day conditions. PMID: 23857347
- COP1 plays a role in setting the daily patterns of sensitivity to shade in the fluctuating light environments of plant canopies. PMID: 23647163
- A functional connection between COP1 and the TOPOVI in plants links COP1-dependent development with the regulation of endoreduplication. PMID: 23573936
- The COP1/SPA complex affects PAP1 and PAP2 both transcriptionally and post-translationally. The COP1/SPA complex controls anthocyanin levels in Arabidopsis. PMID: 23425305
- UV-B-induced reorganization of COP1 complexes achieves a functional switch of COP1 from repressing to promoting photomorphogenesis. PMID: 24067658
- the light-induced decline of phyA levels is reduced in spa mutants irrespective of the growth medium, suggesting a COP1-independent role for SUppressor of phytochrome A proteins. PMID: 23391578
- In cop1 mutants, ddb1b-2 enhanced the cop1-4 short hypocotyl phenotype in dark and light, enhanced anthocyanin levels in cop1-1 in the light, but had no effect in adult plants. PMID: 23450167
- Ala mutants of these tryptophans appear monomeric and constitutively bind COP1 in plants, but their responses indicate that monomer formation and COP1 binding are not sufficient for UVR8 function. PMID: 23012433
- COP1 gene expression in response to photomorphogenic UV-B is controlled by a combinatorial regulation of FHY3 and HY5, and this UV-B-specific working mode of FHY3 and HY5 is distinct from that in far-red light and circadian conditions. PMID: 23150635
- COP1/SPA activity, via LONG HYPOCOTYL IN FR LIGHT1, is required for shade-induced modulation of the auxin biosynthesis pathway and thereby enhances cell elongation in low red:far red. PMID: 23093358
- Ethylene-promoting hypocotyl via IAA is mediated by light, and COP1 has a significant impact on the transcription of some genes downstream of EIN3. Thus, COP1 plays a crucial role in the opposite effects of ethylene on hypocotyl elongation. PMID: 22890836
- Mediator component cooperates with COP1 in the regulation of light responses and that the hypersensitive seedling phenotype strictly depends on the presence of HY5, an important positive regulator of light-dependent gene expression. PMID: 22760208
- COP1 mediates the coordination of root and shoot growth by light through modulation of PIN1- and PIN2-dependent auxin transport in Arabidopsis PMID: 22912415
- COP1 interacts with BBX24 in vivo in a UV-B-dependent manner, and this requires UV-B-induced COP1 accumulation. PMID: 22410790
- Mutations in the region responsible for the interaction with COP1 revealed that a physical interaction of the proteins is also required for degradation of BBX24 in the light and for normal photomorphogenesis. PMID: 21685177
顯示更多
收起更多
-
亞細胞定位:Nucleus. Cytoplasm.
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
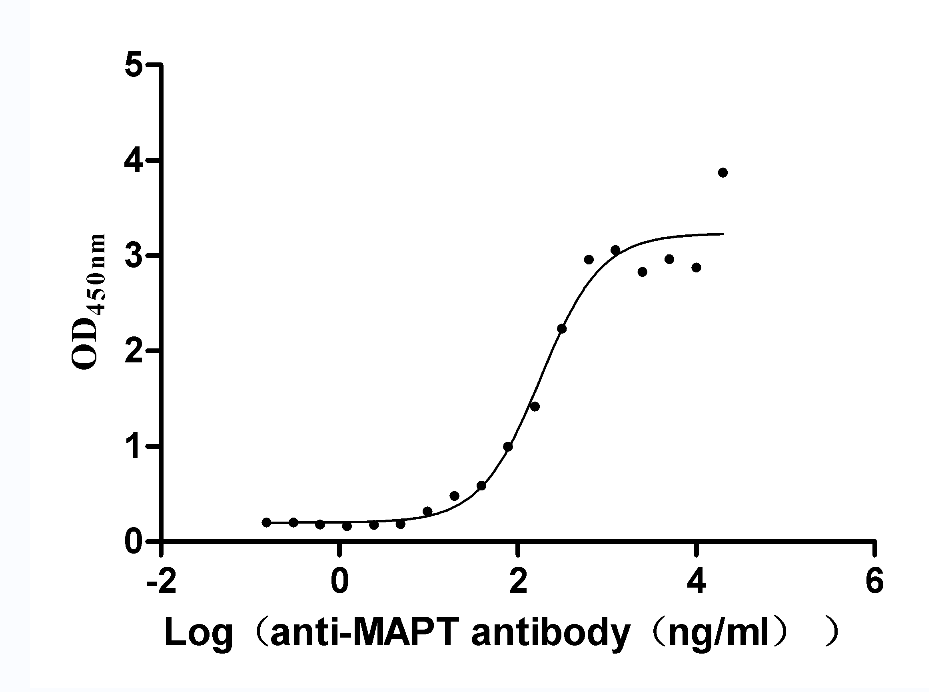
Recombinant Rat Microtubule-associated protein tau (Mapt) (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
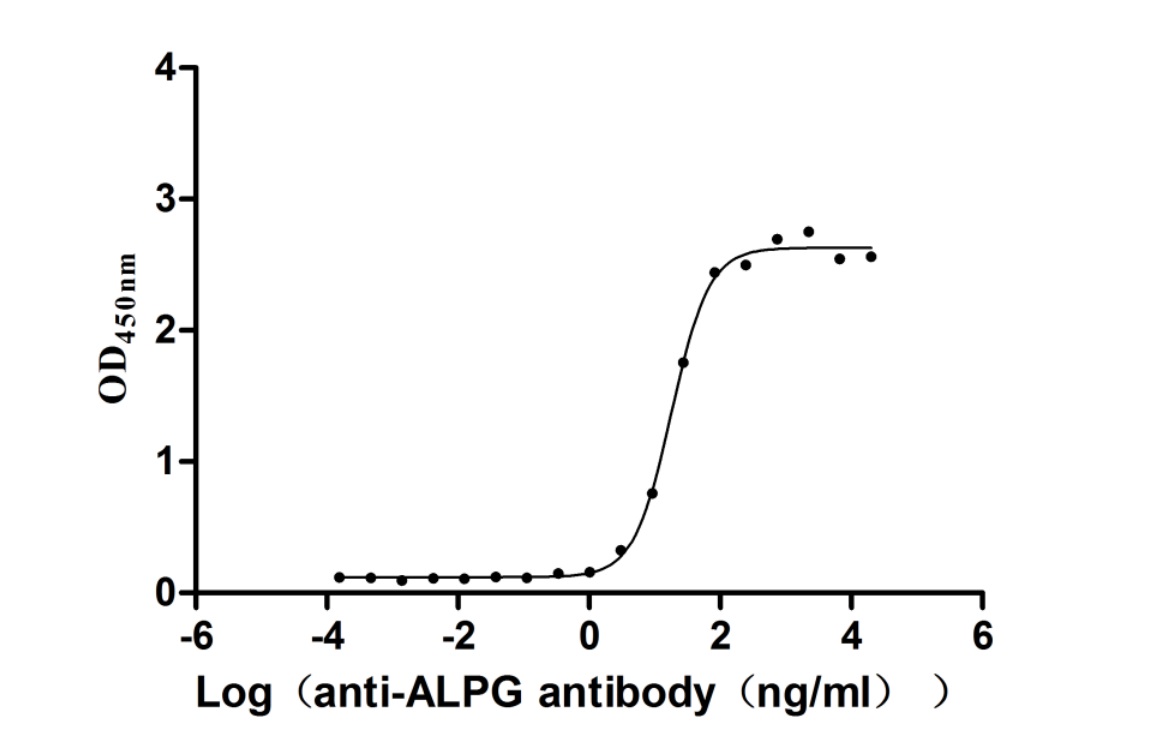
Recombinant Human Alkaline phosphatase, germ cell type (ALPG) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
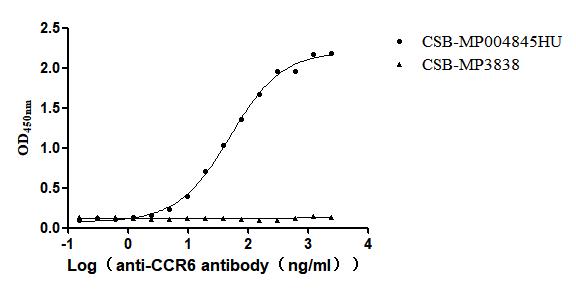
Recombinant Human C-C chemokine receptor type 6(CCR6)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
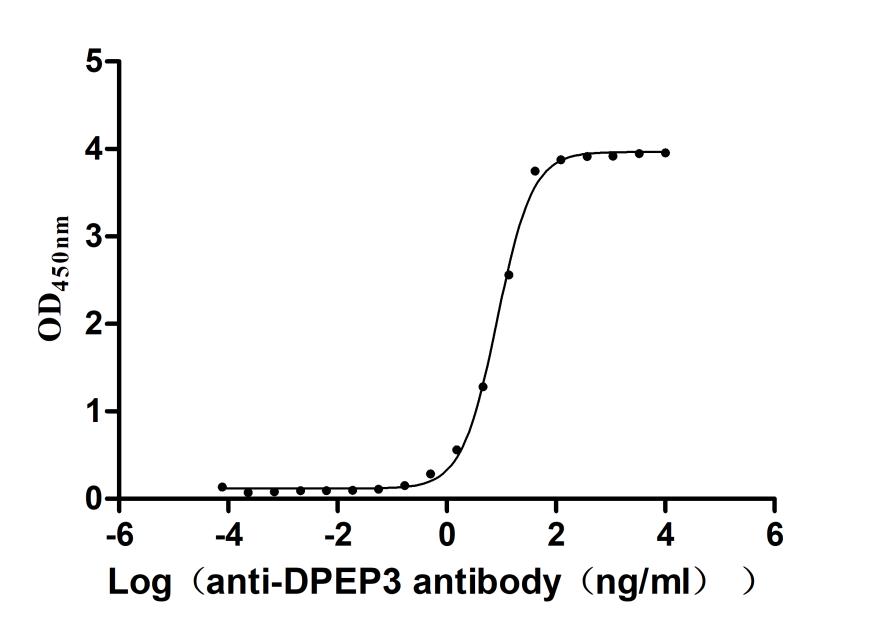
Recombinant Macaca fascicularis Dipeptidase 3(DPEP3) (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant DT3C (Diphtheria toxin & spg 3C domain) for Antibody Internalization Assay (Active)
Express system: E.coli
Species: N/A









