Phospho-HDAC8 (Ser39) Antibody
-
中文名稱:磷酸化-HDAC8 (Ser39)兔多克隆抗體
-
貨號:CSB-PA795062
-
規格:¥2454
-
圖片:
-
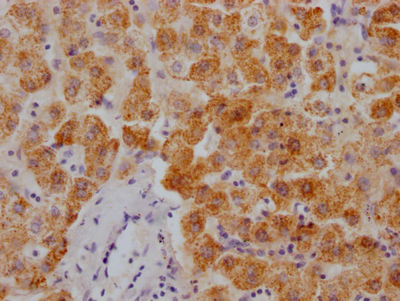
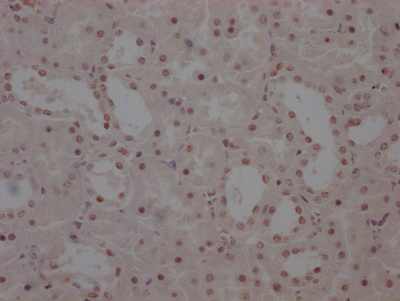
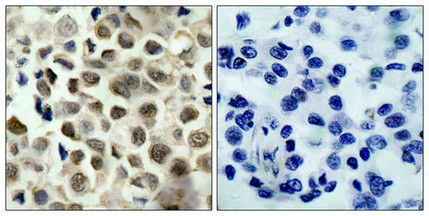 Immunohistochemical analysis of paraffin-embedded human lung carcinoma tissue using HDAC8(Phospho-Ser39) Antibody(left) or the same antibody preincubated with blocking peptide(right).
Immunohistochemical analysis of paraffin-embedded human lung carcinoma tissue using HDAC8(Phospho-Ser39) Antibody(left) or the same antibody preincubated with blocking peptide(right). -
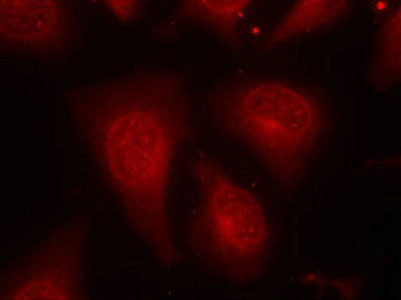 Immunofluorescence staining of methanol-fixed Hela cells using HDAC8(Phospho-Ser39) Antibody.
Immunofluorescence staining of methanol-fixed Hela cells using HDAC8(Phospho-Ser39) Antibody.
-
-
其他:
產品詳情
-
產品名稱:Rabbit anti-Homo sapiens (Human) HDAC8 Polyclonal antibody
-
Uniprot No.:
-
基因名:
-
宿主:Rabbit
-
反應種屬:Human,Mouse,Rat
-
免疫原:Peptide sequence around phosphorylation site of serine 39 (R-A-S(p)-M-V) derived from Human HDAC8.
-
免疫原種屬:Homo sapiens (Human)
-
克隆類型:Polyclonal
-
純化方式:Antibodies were produced by immunizing rabbits with synthetic phosphopeptide and KLH conjugates. Antibodies were purified by affinity-chromatography using epitope-specific phosphopeptide. Non-phospho specific antibodies were removed by chromatogramphy usi
-
濃度:It differs from different batches. Please contact us to confirm it.
-
產品提供形式:Liquid
-
應用范圍:ELISA,IHC,IF
-
推薦稀釋比:
Application Recommended Dilution IHC 1:50-1:100 IF 1:100-1:200 -
Protocols:
-
儲存條件:Upon receipt, store at -20°C or -80°C. Avoid repeated freeze.
-
貨期:Basically, we can dispatch the products out in 1-3 working days after receiving your orders. Delivery time maybe differs from different purchasing way or location, please kindly consult your local distributors for specific delivery time.
-
用途:For Research Use Only. Not for use in diagnostic or therapeutic procedures.
產品評價

相關產品
靶點詳情
-
功能:Responsible for the deacetylation of lysine residues on the N-terminal part of the core histones (H2A, H2B, H3 and H4). Histone deacetylation gives a tag for epigenetic repression and plays an important role in transcriptional regulation, cell cycle progression and developmental events. Histone deacetylases act via the formation of large multiprotein complexes. Also involved in the deacetylation of cohesin complex protein SMC3 regulating release of cohesin complexes from chromatin. May play a role in smooth muscle cell contractility.
-
基因功能參考文獻:
- we report a de novo missense HDAC8 mutation that causes overlapping features of Rett-related disorder and Cornelia de Lange syndrome. It is possible that this HDAC8 mutation may be harmful via MECP recruitment, and may consequently cause Rett-related disorder PMID: 29519750
- this study identifies additional underlying causes of CdLS, describes the first instance of a somatic HDAC8 mutation in an individual with CdLS features and provides insight into the molecular bases of HDAC8 deletions. PMID: 29279609
- The present study identified a novel missense mutation (c.806T>G, p.I269R) in the HDAC8 gene leading to Cornelia de Lange Syndrome (CdLS), which not only provided strong evidence for diagnosis in this present patient, but also expanded the spectrum of pathogenic mutations for CdLS. PMID: 29991052
- chemical crosslinking approach identified novel HDAC8-specific substrates with high catalytic efficiency, thus presenting a general strategy for unbiased deacetylase substrate discovery PMID: 29035536
- Co2+ and Zn2+ are the most active metals in HDAC8 biocatalysis, followed by Fe2+, and Mn2+ and Mg2+ to be the least active. PMID: 26996235
- MSC from MPN have higher expression of HDAC8 than normal ones, and the inhibition of HDAC8 expression by its specific inhibitor decreases the capacity of the stroma to support hematopoietic cells from MPN patients, suggesting that HDAC8 may be a potential therapeutic target in this setting. PMID: 28390197
- This study for the first time demonstrated that HDAC8 activity determines susceptibility to cell cycle arrest induced by Anthrax Lethal Toxin, through regulating the PI3K-PTEN-AKT signaling axis. PMID: 28509866
- AHR regulated cell proliferation and tumorigenesis by directly targeting and activating HDAC8 expression in hepatocellular carcinoma cells. PMID: 27283490
- The Epac-Rap1-Akt pathway mediates cAMP signaling-induced inhibition of JNK-dependent HDAC8 degradation, and the resulting HDAC8 increase augments cisplatin-induced apoptosis by repressing TIPRL expression in H1299 lung cancer cells. PMID: 28232663
- The X-ray crystal structure of HDAC8 complexed with inhibitor confirms the formation of a unique subpocket in HDAC8. PMID: 27374062
- These data provide support for the hypothesis that HDAC8 may undergo metal switching in vivo that, in turn, may regulate its activity. However, future studies are needed to explore the identity of the metal ion bound to HDAC8 in cells under varied conditions PMID: 28937750
- The study reports the crystal structure of the HDAC8-trapoxin A complex at 1.24 A resolution, revealing that the ketone moiety of l-Aoe undergoes nucleophilic attack to form a zinc-bound tetrahedral gem-diolate that mimics the tetrahedral intermediate and its flanking transition states in catalysis. PMID: 28846375
- The findings suggest that miR-455-3p plays a critical role during chondrogenesis by directly targeting HDAC2/8 and promoting histone H3 acetylation. PMID: 27638301
- 7-amino-4-methylcoumarin did not affect acetyllysine preference in a multiply acetylated substrate. In contrast, AMC significantly enhanced KDAC6 substrate affinity, greatly reduced Sirt1 activity, eliminated the substrate sequence specificity of KDAC4, and had no consistent effect with KDAC8 substrates. PMID: 28749131
- Enzymological, biophysical, and structural studies of Cornelia de Lange Syndrome HDAC8 protein mutants have yielded critical insight on compromised catalysis in vitro. Most CdLS HDAC8 mutations trigger structural changes that directly or indirectly impact substrate binding and catalysis. PMID: 27576763
- class I HDACs (HDAC1, 2, 3 and 8) play a major role in regulating extracellular matrix and Epithelial-mesenchymal transition, whereas class IIa HDACs (HDAC4 and 5) are less effective. PMID: 27420561
- Losses of catalytic efficiency in histone deacetylase 8 (HDAC8) are observed for G304A and G305A mutations. PMID: 27933794
- Data suggest sequencing of histone deacetylase 8 protein (HDAC8) as an indispensable part of the routine molecular diagnostic for patients with Cornelia de Lange syndrome (CdLS) or CdLS-overlapping features. PMID: 26671848
- HDAC8 were overexpressed in oral squamous cell carcinoma tissues, mainly localized in the cytoplasm. PMID: 28004115
- Study reveals that HDAC8 can bind and catalyze deacetylation of many acetylated peptides with sequences corresponding to cellular, non-histone proteins, thereby opening a new window to the functional role of HDAC8 in cells. PMID: 26933971
- Findings suggest the therapeutic potential of histone deacetylase 8 histone (HDAC8)inhibition to suppress Notch1 signaling in breast cancer. PMID: 26625202
- Data show that histone deacetylase 8 (HDAC8) inhibition led to accumulation of acetylated-SMC3 protein but had no influence on the transcription of estrogen-responsive genes. PMID: 27072133
- Results provide direct evidence that potential KDAC8 substrates can be directly deacetylated by KDAC8. PMID: 26402585
- study elucidates an HDAC8-mediated p53-inactivating mechanism promoting leukemia stem cells activity PMID: 26387755
- Studies indicate that histone deacetylase 8 protein (HDAC8) aberrantly deacetylates p53 protein and promotes leukemia stem cells (LSCs) transformation and maintenance. PMID: 26923266
- Our data exhibited an important role of HDAC8 in promoting gastric cancer tumorigenesis and identify this HDAC8 as a potential therapeutic target for the treatment of gastric cancer. PMID: 26412386
- The H143A and H142A/H143A mutants exhibit activity that is >80000-fold lower than that of wild-type HDAC8; the buried D176N and D176A mutants have significant catalytic effects, with more subtle effects caused by D183N and D183A PMID: 26806311
- cAMP signaling increases HDAC8 protein levels by reducing JNK-mediated autophagy and ubiquitin-proteasome-dependent degradation of the HDAC8 protein in H1299 lung cancer cells. PMID: 26792731
- HDAC8i induced cell growth inhibition and marked S-phase cell cycle arrest in human and murine-derived MPNST cells PMID: 26200462
- Report the preparation and biophysical evaluation of five HDAC8 mutants: P91L, G117E, H180R, D233G, and G304R. Additionally, the double mutants D233G-Y306F and P91L-Y306F were prepared to enable cocrystallization of intact enzyme-substrate complexes. PMID: 26463496
- HDAC8 was increased in BRAF-mutated melanoma. Increased cytoplasmic HDAC8 immunoreactivity was independently associated with an improved survival from both diagnosis of primary melanoma and from first detection of stage IV disease to melanoma death PMID: 25836739
- findings show how HDAC8 drives nonalcoholic fatty liver disease-associated hepatocarcinogenesis PMID: 26383163
- Data reveal a role for miR-21-3p in regulating HDAC8 expression and Akt/Gsk3beta pathway in cardiac hypertrophy. PMID: 25504627
- Studied the kinetics, thermodynamics, and selectivity of Zn(II) and Fe(II) binding to HDAC8. PMID: 25516458
- HDAC8 X-ray crystal structures reveal that each Cornelia de Lange syndrome mutation causes local structural changes that compromise catalysis and/or thermostability. PMID: 25075551
- report the X-ray crystal structures of HDAC8 complexed with three synthetic analogues of Largazole in which the depsipeptide ester is replaced with a rigid amide linkage PMID: 25793284
- On the corresponding BMF gene promoter, loss of HDAC8 was associated with signal transducer and activator of transcription 3 (STAT3)/specificity protein 3 (Sp3) transcription factor exchange and recruitment of p300. PMID: 25321483
- Our mechanistic and cellular studies on HDAC8 activation have the potential to provide insight into the development of novel anticancer drugs. PMID: 25605725
- Deregulation of HDAC8 is frequent in urothelial cancer, but neither specific pharmacological inhibition nor siRNA-mediated knockdown of HDAC8 impaired viability of urothelial cancer cell lines in a therapeutic useful manner. PMID: 25011684
- histone deacetylase 8 inhibition reduces gene expression and production of proinflammatory cytokines in vitro and in vivo PMID: 25451941
- detailed thermodynamic studies of the binding of structurally similar ligands, which differed with respect to the "cap", "linker", and "metal-binding" regions of the suberoylanilide hydroxamic acid pharmacophore, to HDAC8 PMID: 25407689
- Loss-of-function mutations in HDAC8 cause a range of overlapping phenotypic spectrum of Cornelia de Lange syndrome-like feature. PMID: 24403048
- In response to contractile stimulation, HDAC8 may mediate cortactin deacetylation, which subsequently promotes actin filament polymerization and smooth muscle contraction. PMID: 24920679
- DEC1 coordinates with HDAC8 to differentially regulate TAp73 and DeltaNp73 expression. PMID: 24404147
- Data indicate prominent changes in urothelial cancer cell lines (UCC) were HDAC2 and/or HDAC8 up-regulation. PMID: 22944197
- Using molecular dynamics simulations the study found a mechanism whereby the interactions and dynamics of two loops tune the configuration of functionally important residues of HDAC8 and could therefore influence the activity of the enzyme. PMID: 24171457
- HDAC8 isoverexpressed in hepatocellular carcinoma; knockdown suppresses tumor growth and enhances apoptosis PMID: 24077923
- HDAC8 plays an important role in the modulation of SOCS1 and SOCS3 by curcumin. PMID: 23430957
- SOX4 is a direct target gene of FRA-2 and induces expression of HDAC8 in adult T-cell leukemia/lymphoma. PMID: 23482931
- Data indicate that knockdown of HDAC8 resulted in the increased expression of SOCS1 and SOCS3, and overexpression of SOCS1 and SOCS3 significantly inhibited cell growth and suppressed JAK2/STAT signaling. PMID: 23111066
顯示更多
收起更多
-
相關疾病:Cornelia de Lange syndrome 5 (CDLS5); Wilson-Turner X-linked mental retardation syndrome (WTS)
-
亞細胞定位:Nucleus. Cytoplasm. Note=Excluded from the nucleoli. Found in the cytoplasm of cells showing smooth muscle differentiation.
-
蛋白家族:Histone deacetylase family, HD type 1 subfamily
-
組織特異性:Weakly expressed in most tissues. Expressed at higher level in heart, brain, kidney and pancreas and also in liver, lung, placenta, prostate and kidney.
-
數據庫鏈接:
Most popular with customers
-
-
YWHAB Recombinant Monoclonal Antibody
Applications: ELISA, WB, IHC, IF, FC
Species Reactivity: Human, Mouse, Rat
-
Phospho-YAP1 (S127) Recombinant Monoclonal Antibody
Applications: ELISA, WB, IHC
Species Reactivity: Human
-
-
-
-
-