Recombinant Mouse Interleukin-5 protein (Il5)
In Stock-
中文名稱:小鼠Il5重組蛋白
-
貨號:CSB-AP003311MO
-
規(guī)格:¥852
-
圖片:
-
-
其他:

產(chǎn)品詳情
-
純度:>98% as determined by SDS-PAGE.
-
內(nèi)毒素:Less than 1.0 EU/μg as determined by LAL method.
-
生物活性:Fully biologically active when compared to standard. The ED50 as determined by a cell proliferation assay using human TF-1 cells is less than 2 ng/ml, corresponding to a specific activity of >5.0x105 IU/mg.
-
基因名:
-
Uniprot No.:
-
別名:Il5; Il-5Interleukin-5; IL-5; B-cell growth factor II; BCGF-II; Cytotoxic T-lymphocyte inducer; Eosinophil differentiation factor; T-cell replacing factor; TRF
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length of Mature Protein
-
來源:E.Coli
-
分子量:13.1 kDa
-
表達(dá)區(qū)域:21-133aa
-
氨基酸序列MEIPMSTVVK ETLTQLSAHR ALLTSNETMR LPVPTHKNHQ LCIGEIFQGL DILKNQTVRG GTVEMLFQNL SLIKKYIDRQ KEKCGEERRR TRQFLDYLQE FLGVMSTEWA MEG
-
蛋白標(biāo)簽:Tag-Free
-
產(chǎn)品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:0.2 m filtered 20 mM Tris, pH 9.0, 150 mM NaCl ,lyophilized
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:5-10 business days
-
Datasheet & COA:Please contact us to get it.
產(chǎn)品評價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:Factor that induces terminal differentiation of late-developing B-cells to immunoglobulin secreting cells.
-
基因功能參考文獻(xiàn):
- binding of IL-5 to IL-5Ralpha receptors enhances angiogenic responses by stimulating the expression of HSP70-1 via the eNOS signaling pathway. PMID: 28317868
- IL-33 acts directly on bone marrow ILC2s, making them an early source of IL-5 and part of a process that is central in IL-33-driven eosinophilia. PMID: 28921511
- Obesity alters the lung neutrophil infiltration to enhance breast cancer metastasis through IL5 and GM-CSF. PMID: 28737771
- these studies establish a basal defect in eosinophilopoiesis in IL-33- and ST2-deficient mice and a mechanism whereby IL-33 supports eosinophils by driving both systemic IL-5 production and the expansion of IL-5Ralpha-expressing precursor cells PMID: 27683753
- Increased production of IL-5 from Peyer's patch cells and the restored Th1-type immune response might cause the production of abnormal IgA and might induce the glomerular deposition of IgA in IGA nephropathy. PMID: 26719095
- selective proliferation of IgM rheumatoid factor-secreting B-1a cells is induced by co-stimulation by the specific pathogen antigen and IL-5 in the development of MC in Capillaria hepatica-infected mice PMID: 25452118
- IL5, a cytokine involved in allergic and infectious diseases, facilitates metastatic colonization through recruitment of sentinel eosinophils and regulation of other inflammatory/immune cells in the microenvironment of the distal lung. PMID: 25691457
- Data (including data from knockout mice) suggest that up-regulation of IL5 production in lungs during influenza virus infection is due to infiltration of natural killer cells and alveolar macrophages into infected lung tissue. PMID: 24068930
- A decrease in the levels of IL-5, IL-9, and IL-6R in the BALF. PMID: 24246030
- eosinophils express CAR4 following IL-5 or allergen exposure, and that CAR4 is involved in regulating the lung transcriptome associated with allergic airway inflammation PMID: 24808371
- Protection of montelukast on OVA-induced eosinophilic gastroenteritis via modulating IL-5, eotaxin-1 and MBP expression. PMID: 23855447
- Together, these studies support the conclusion that surfactant protein D increases susceptibility to Cryptococcus neoformans infection by promoting Cryptococcus neoformans-driven pulmonary IL-5 and eosinophil infiltration. PMID: 24478083
- IL5 induced eosinophils and cysteinyl leukotrienes are involved in the pathology of mite antigen-induced chronic asthma model. PMID: 23942524
- Id3 is a key regulator of natural helper cell IL-5 production and B-1a B cell homeostasis. PMID: 24115031
- Interleukin-5 plays a key role in mouse strain- dependent susceptibility to contact hypersensitivity through its effects on initiator B cells. PMID: 23711860
- macrophage IL-5 is a target gene for LXR activation, and the induction of macrophage IL-5 expression can be related to LXR-inhibited atherosclerosis. PMID: 23150660
- Ang II induces increased Th2 cytokines IL-5 and IL-10 early in the course of experimental abdominal aortic aneurysm formation, and inhibition of IL-5 prevents AAA formation suggesting an important role. PMID: 22459292
- Sex difference in IL-5 production by splenocytes might be due, at least in part, to the sex difference in the sensitivity of CD4+ T cells to suppression by CD8+ T cells. PMID: 22627364
- Invading tumor cells enhance and increase local IL-5 production from innate IL-5-producing non-T lymphoid cells residing in the intestine, peritoneal cavity and lungs of naive mice. PMID: 22174445
- PARP-1 regulates Il-5 production through calpain degradation of STAT-6 in a murine asthma model. PMID: 21276008
- IL-5 production by splenocytes triggered by TCR activation was higher in female mice than in male mice, and the difference might be attributable to sex differences in CD4+ and CD8+ T cell functions. PMID: 21646791
- Transnasal administration of liposome- mediated IL12 could depress the expression of IL-5 in bone marrow, peripheral blood, and nasal mucosa in allergic rhinitis. PMID: 19954023
- Exacerbation of oxazolone colitis by infection with the helminth Hymenolepis diminuta: involvement of IL-5 and eosinophils. PMID: 21037078
- Greater antigen-induced Th2 IL-5 production by bronchial lymph node cells from female mice was associated with enhanced Th2 cell differentiation and increased expression of the Th2-specific transcription factor, GATA-3. PMID: 20337994
- IL-5 production by bronchial epithelial cells can impact the microenvironment of the lung, modifying pathologic and protective immune responses in the airways PMID: 20494340
- IL-5 promotes eosinophil trafficking to the esophagus PMID: 11859139
- IL-5 is required for the development of tissue and marrow eosinophilia, the formation of eosinophil/basophil colony-forming units, and the early development of symptoms in experimental allergic rhinitis. PMID: 11884474
- Role of IL-5 during primary and secondary immune response to acetylcholine receptor. PMID: 11960640
- mechanism of synergism between eotaxin and IL5, facilitating the selective recruitment of eosinophils into sites of allergic inflammation PMID: 12083417
- A putative Bcl6-binding DNA sequence which acts as a silencer element has been identified in the 3' untranslated region of IL-5 cDNA. PMID: 12097386
- IL-5 alone does not account for the complexities of bronchopulmonary hyperreactivity or of eosinophil tissue trapping PMID: 12231478
- effects of constitutive interleukin-5 (IL-5) expression and overabundance of eosinophils on the development and function of the mammary gland, uterus, and ovary PMID: 12620930
- IL-5 appears to be required for the accumulation of eosinophils and airway hyperresponsiveness in the inflammatory lung. PMID: 12660425
- The ability of IL-13 to induce eosinophilic esophagitis was abolished in STAT6-deficient mice, nearly completely ablated in IL-5-deficient mice, and significantly diminished in eotaxin-1-deficient mice. PMID: 14598258
- immune effector mechanisms in murine filarial infection are dependent on both IFN-gamma and IL-5, whose synergistic effects may be mediated, at least in part, by neutrophils for the control of adult worms. PMID: 14638787
- These results suggest an important role for interleukin-5, eosinophils, alphaVbeta6 integrin, and TGF-beta in airway remodeling. PMID: 14966564
- Pulmonary fibrosis lesions are abolished in sensitized and allergen-exposed IL-5 receptor-null mice, whereas they are markedly accentuated in IL-5 transgenic animals. PMID: 14975941
- Marked impairment of the maintenance of mature B-1 lymphocyte survival and homeostatic proliferation is demonstrated by blocking IL-5 signals. The key question is to what extent IL-5 is involved in mature B-1 cell survival and homeostatic proliferation. PMID: 15128785
- IL-5 participates in the pathogenesis of ileitis in SAMP1/Yit mice. PMID: 15162425
- CD4(-)c-kit(-)CD3epsilon(-)IL-2Ralpha(+) Peyer's patch cells are capable of secreting a high level of IL-5 in response to IL-2 PMID: 15214040
- IL-5 links adaptive and natural immunity for epitopes of oxidized LHDL and protects from arteriosclerosis. PMID: 15286809
- Anti-IL-5 was able to reduce eosinophil numbers in all tissue compartments, as well as BrdU+ eosinophils and CD34+ progenitor cells, and in all instances to a greater extent than anti-IL-9. PMID: 15823208
- the increase in airway eosinophilia seen with COX inhibition is dependent on IL-5, whereas the increase in airway hyperresponsiveness is not PMID: 16339565
- IL-5 is not necessary for differential splicing to occur in vivo, as all three forms of the IL-5R alpha are detected in both strains of IL-5 gene-deleted mice PMID: 16856933
- It was found that MCA-induced tumor incidence and growth were significantly attenuated in IL-5 transgenic mice. PMID: 17371978
- Il-5 level peaked at 7 dpi in bronchoalveolar lavage fluid. PMID: 17487773
- IL-5 gene delivery suppresses sensitization to antigen (ovalbumin) by upregulating transforming growth factor beta 1-dependent signaling to CD4-expressing T cells, thus suppressing allergic airway inflammation. PMID: 17579048
- cyclic AMP signals enhance histone H3 acetylation at the IL-5 promoter and the concerted binding of GATA-3 and NFATc to the promoter. PMID: 18772129
- there is a reciprocal relationship between inducible nitric oxide synthase and poly(ADP-ribose) polymerase-1; expression of inducible nitric oxide synthase may be dispensable for eosinophilia after interleukin-5 production PMID: 18829681
- Healing was significantly delayed in IL-5-overexpressing mice with wounds gaping wider and exhibiting impaired re-epithelialization PMID: 18839016
顯示更多
收起更多
-
亞細(xì)胞定位:Secreted.
-
蛋白家族:IL-5 family
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
-AC1.jpg)
Recombinant Human HLA class II histocompatibility antigen gamma chain (CD74), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
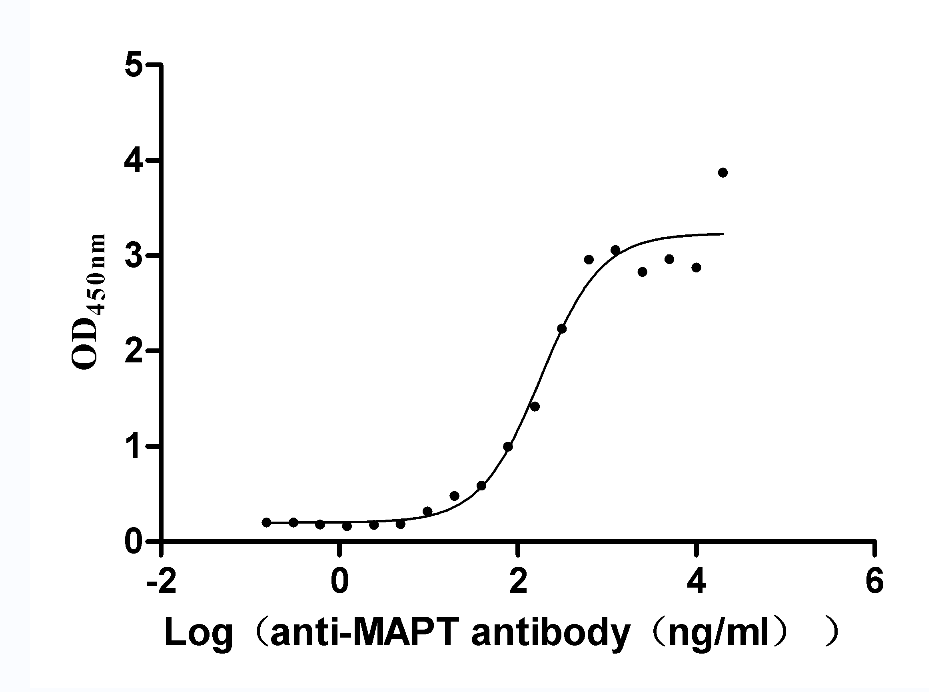
Recombinant Rat Microtubule-associated protein tau (Mapt) (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
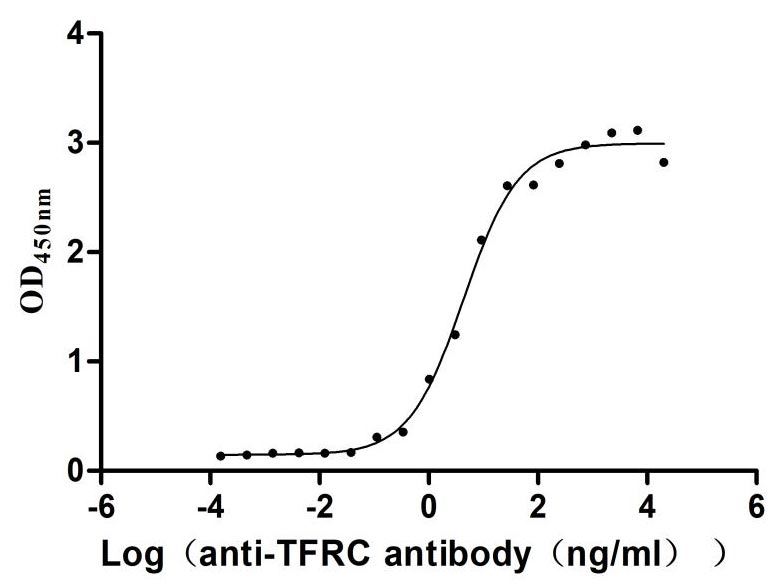
Recombinant Human Transferrin receptor protein 1 (TFRC), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
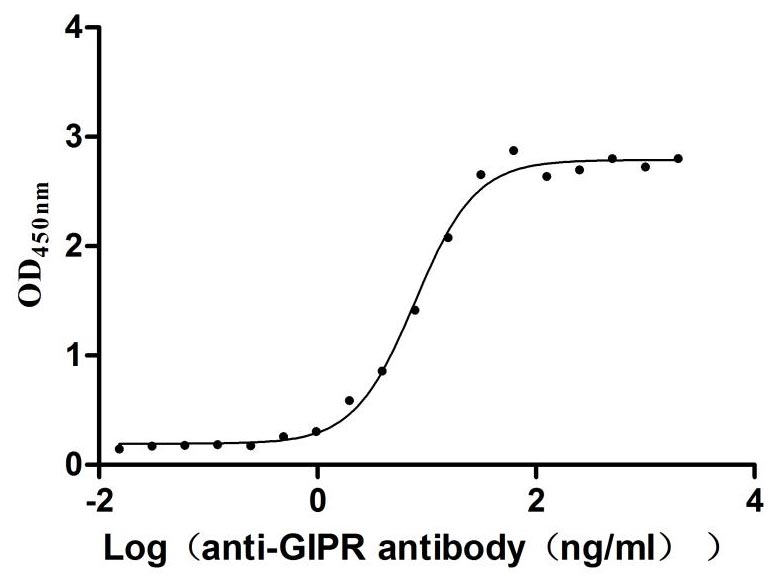
Recombinant Rat Gastric inhibitory polypeptide receptor (Gipr), partial (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
Recombinant Human Urokinase-type plasminogen activator(PLAU) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
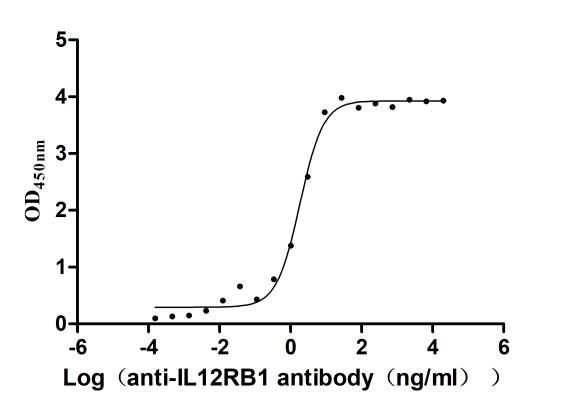
Recombinant Human Interleukin-12 receptor subunit beta-1(IL12RB1),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Human Tumor necrosis factor ligand superfamily member 15(TNFSF15) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
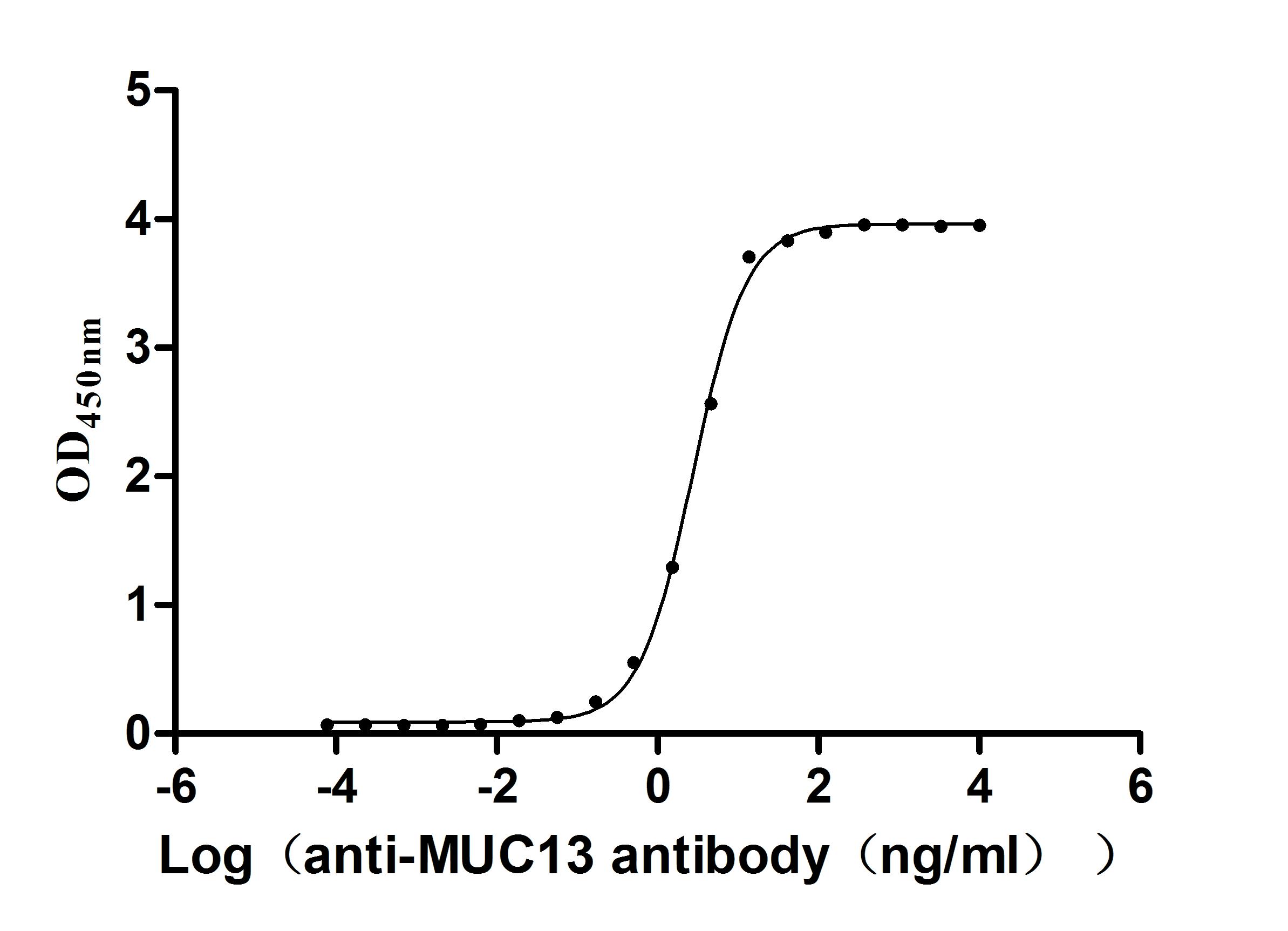
Recombinant Human Mucin-13(MUC13),partial (Active)
Express system: yeast
Species: Homo sapiens (Human)









