-
中文名稱:小鼠脂蛋白脂酶(LPL)酶聯免疫試劑盒
-
貨號:CSB-E08495m
-
規格:96T/48T
-
價格:¥3800/¥2500
-
其他:

產品詳情
-
產品描述:小鼠脂蛋白脂酶(LPL)酶聯免疫試劑盒(CSB-E08495m)為雙抗夾心法ELISA試劑盒,定量檢測血清、血漿、組織培養上清液、組織勻漿樣本中的LPL含量。LPL即脂蛋白脂肪酶,是脂肪代謝關鍵酶。它在血漿脂蛋白代謝、能量平衡等過程中發揮重要作用。研究機制方面,其能水解乳糜微粒和極低密度脂蛋白中的甘油三酯,產生游離脂肪酸供組織攝取利用,在脂代謝疾病等研究中備受關注。試劑盒檢測范圍為0.625 ng/mL-40 ng/mL,適用于小鼠模型中脂代謝調控機制研究、心血管疾病模型評估及藥物干預實驗等科研場景本品僅用于科研,不用于臨床診斷,產品具體參數及操作步驟詳見產品說明書。
-
別名:Lpl ELISA Kit; Lipoprotein lipase ELISA Kit; LPL ELISA Kit; EC 3.1.1.34 ELISA Kit
-
縮寫:
-
Uniprot No.:
-
種屬:Mus musculus (Mouse)
-
樣本類型:serum, plasma, cell culture supernates, tissue homogenates
-
檢測范圍:0.625 ng/mL-40 ng/mL
-
靈敏度:0.156 ng/mL
-
反應時間:1-5h
-
樣本體積:50-100ul
-
檢測波長:450 nm
-
研究領域:Metabolism
-
測定原理:quantitative
-
測定方法:Sandwich
-
精密度:
Intra-assay Precision (Precision within an assay): CV%<8% Three samples of known concentration were tested twenty times on one plate to assess. Inter-assay Precision (Precision between assays): CV%<10% Three samples of known concentration were tested in twenty assays to assess. -
線性度:
To assess the linearity of the assay, samples were spiked with high concentrations of mouse LPL in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay. Sample Serum(n=4) 1:1 Average % 91 Range % 87-93 1:2 Average % 92 Range % 88-95 1:4 Average % 94 Range % 90-97 1:8 Average % 100 Range % 95-103 -
回收率:
The recovery of mouse LPL spiked to levels throughout the range of the assay in various matrices was evaluated. Samples were diluted prior to assay as directed in the Sample Preparation section. Sample Type Average % Recovery Range Serum (n=5) 94 91-97 EDTA plasma (n=4) 95 92-98 -
標準曲線:
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed. 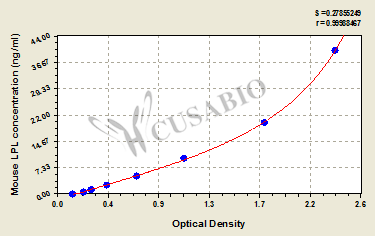
ng/ml OD1 OD2 Average Corrected 40 2.409 2.348 2.379 2.233 20 1.788 1.767 1.778 1.632 10 1.102 1.088 1.095 0.949 5 0.691 0.688 0.690 0.544 2.5 0.444 0.434 0.439 0.293 1.25 0.301 0.308 0.305 0.159 0.625 0.232 0.242 0.237 0.091 0 0.144 0.148 0.146 -
數據處理:
-
貨期:3-5 working days
引用文獻
- Gut Bifidobacterium pseudocatenulatum protects against fat deposition by enhancing secondary bile acid biosynthesis A Zha, M Qi, Y Deng, H Li, N Wang, C Wang, S Liao,iMeta,2024
- Honokiol Prevents Intestinal Barrier Dysfunction in Mice with Severe Acute Pancreatitis and Inhibits the JAK/STAT1 Pathway and Acetylation of HMGB1 J Li,/,2023
- Maternal malnutrition impacts placental morphology and transporter expression: an origin for poor offspring growth Connor KL,J Nutr Biochem,2020
- Maternal malnutrition impacts placental morphology and transport. An origin for poor offspring growth and vulnerability to disease Kristin L Connor, et al,Biorxiv,2019
- Changes of vitellogenin and Lipase in captive Sterlet sturgeon Acipenser ruthenus females during previtellogenesis to early atresia. Sobhan Akhavan.et al,Fish Physiology and Biochemistry,2016
- NMDA Receptor Antagonist Attenuates Bleomycin-Induced Acute Lung Injury. Yang Li, et al,PLoS One,2015
- Effect of yellow capsicum extract on proliferation and differentiation of 3T3-L1 preadipocytes Feng Z et al,Nutrition,2013
產品評價

相關產品
相關問答
■ 常見問題解答
Q:
1) Concentration of sodium azide or thimerosal and 2-mercaptoethanol(or other toxic substances) if it is contained in the product.
2) Please let us know whether the CSB-E08495m was produced with Baculovirus to
follow Cartagena protocol regulation.
A:
Thanks for your inquiry.
CSB-E08495m,
The stop solution is 4N sulfuric acid.
There is no sodium azide or thimerosal and 2-mercaptoethanol(or other toxic substances) in the kit.
The standard is recombinant protein expressed in the eukaryotic system. There is no Baculovirus derived component.
CSB-E08495m,
The stop solution is 4N sulfuric acid.
There is no sodium azide or thimerosal and 2-mercaptoethanol(or other toxic substances) in the kit.
The standard is recombinant protein expressed in the eukaryotic system. There is no Baculovirus derived component.
靶點詳情
-
功能:Key enzyme in triglyceride metabolism. Catalyzes the hydrolysis of triglycerides from circulating chylomicrons and very low density lipoproteins (VLDL), and thereby plays an important role in lipid clearance from the blood stream, lipid utilization and storage. Although it has both phospholipase and triglyceride lipase activities it is primarily a triglyceride lipase with low but detectable phospholipase activity. Mediates margination of triglyceride-rich lipoprotein particles in capillaries. Recruited to its site of action on vascular endothelium by binding to GPIHBP1 and cell surface heparan sulfate proteoglycans.
-
基因功能參考文獻:
- LPL-mediated release of essential fatty acid DHA regulates hematopoietic stem progenitor cell expansion and definitive hematopoiesis PMID: 29615667
- the negatively charged IDR of GPIHBP1 traverses a vast space, facilitating capture of LPL by capillary endothelial cells and simultaneously contributing to GPIHBP1's ability to preserve LPL structure and activity. PMID: 29899144
- LPL in the hypothalamus is an important regulator of body weight and glucose homeostasis PMID: 28456865
- These results identify LPL as an important regulator of fatty acid transport to skeletal compartments and demonstrate an intricate functional link between systemic and skeletal fatty acid and glucose metabolism. PMID: 28608812
- mutation of a conserved cysteine in GPIHBP1 abolishes the ability of GPIHBP1 to bind LPL PMID: 28476858
- The data suggests that ANGPTL3 is part of the machinery causing dyslipidemia majorily via LPL inhibition in mastitis mice. PMID: 29104012
- Using in vitro ketosis model by glucose starvation, studied inhibition of ketosis by momilactone B. Found momilactone B could regulate the angiopoietin-like-3 (ANGPTL3)-lipoprotein lipase (LPL)pathway, and suppressed the expression of HMGCS2 through the increased expression of STAT5b. PMID: 27874312
- physiological changes in adipose tissue ANGPTL4 expression during fasting and cold resulted in inverse changes in the amount of mature-glycosylated LPL in wild-type mice, but not Angptl4(-/-) mice. We conclude that ANGPTL4 promotes loss of intracellular LPL by stimulating LPL degradation after LPL processing in the endoplasmic reticulum (ER). PMID: 27034464
- LPL moved quickly from heparan sulfate proteoglycans (HSPGs) on adipocytes to GPIHBP1-coated beads, thereby depleting LPL stores on the surface of adipocytes. We conclude that HSPG-bound LPL in the interstitial spaces of tissues is mobile, allowing the LPL to move to GPIHBP1 on endothelial cells PMID: 27811232
- our study reveals that hepatic LPL is involved in the regulation of plasma LPL activity and lipid homeostasis. PMID: 27234787
- The induction of LPL activity by fasting in core transgenic mice activated PPARalpha downstream target genes that are involved in fatty acid beta-oxidation. PMID: 27665576
- This study shows that TNF-alpha, by a Foxo1 dependent pathway, increases the transcription of ANGPTL4 which is secreted by the cells and causes inactivation of LPL. PMID: 28215713
- Our findings suggest that neuronal LPL is involved in the regulation of body weight and composition in response to either the change in quantity (HF feeding) or quality (n-3 PUFA-enriched) of dietary fat PMID: 27282869
- An LPL structural model suggests that the LPL S447X truncation exposes residues implicated in LPL binding to lipoprotein binding uptake receptors, such as GPIHBP1. PMID: 27984852
- feeding induces lipasin, activating the lipasin-Angptl3 pathway, which inhibits LPL in cardiac and skeletal muscles to direct circulating TAG to WAT for storage PMID: 26687026
- MiR-590 agomir down-regulates LPL mRNA and protein expression in a mouse model of atherosclerosis. PMID: 26397958
- Deficiency of Lipoprotein Lipase in Neurons Decreases AMPA Receptor Phosphorylation and Leads to Neurobehavioral Abnormalities in Mice PMID: 26263173
- Systemic LPL deletion results in impaired glucose tolerance, whole-body and tissue-specific insulin resistance, which is associated with tissue lipid deposition in various insulin target tissues PMID: 25635326
- Results indicated that aggregation of alpha-syn and reduction of UCHL1 expression in LPL-deficient mice may affect synaptic function. PMID: 25595992
- the amount of LPL expressed in muscle and heart governed both the binding of chylomicron particles and the assimilation of chylomicron lipids in the tissue. PMID: 25589507
- Maternal overnutrition induces LPL expression in trophoblasts by reducing the inhibitory effect of SIRT1 on PPARgamma. PMID: 25948680
- Lipoprotein lipase is an important modulator of lipid uptake and storage in hypothalamic neurons. PMID: 26265042
- Results suggest that impaired synaptic vesicle recycling results from deficient docosahexaenoic acid and arachidonic acid and contributes to the presynaptic dysfunction and plasticity impairment in LPL-deficient neurons PMID: 25194787
- Adipocyte-specific Sel1L-deficient (AKO) mice are resistant to diet-induced obesity. Sel1L stabilizes and prevents LPL dimers from aggregation in the endoplasmic reticulum. PMID: 25066055
- This study showed that phloridzin improved plasma lipoprotein lipase activity via a decrease of ANGPTL4 mRNA expression and an increase of AMP-activated protein kinase phosphorylation. PMID: 24932810
- TRL margination depends on LPL bound to GPIHBP1. PMID: 24726386
- LpL hydrolysis of circulating lipoproteins is required for the accumulation of lipids in the heart of fasting mice. PMID: 24493834
- The expression levels of miR-27a and miR-29a inversely correlate with the mRNA levels of lipoprotein lipase and its key transcriptional regulator peroxisome proliferator-activated receptor gamma during 3T3-L1 adipocyte differentiation. PMID: 24457907
- Leu452His mutation in lipoprotein lipase gene transfer associated with hypertriglyceridemia in mice in vivo. PMID: 24086538
- PPARgamma1 was intimately involved in LPL gene expression in skeletal muscle and the AMPK-PPARgamma1 pathway may play a role in exercise-induced LPL expression. PMID: 24644240
- Activated cyclin-dependent kinase 5 promotes microglial phagocytosis of fibrillar beta-amyloid by up-regulating lipoprotein lipase expression. PMID: 23816988
- inactivation of LPL by Angptl4 appears to occur after both proteins have traveled along the secretory pathway and arrived at the cell surface. PMID: 24220340
- The present study showed that miR-467b protects apoE(-/-) mice from atherosclerosis by reducing lipid accumulation and inflammatory cytokine secretion via downregulation of LPL expression. PMID: 24309104
- Lipoprotein lipase activity decreases in adipose tissue during fasting. PMID: 23176178
- Macrophage LpL plays an important role in the development of atherosclerosis but not adiposity. PMID: 23378601
- LpL has a role as the "gatekeeper" for tissue lipid distribution and its deficiency more profoundly affects brown than white fat biology PMID: 23542081
- Neither a high fat diet nor fasting/re-feeding markedly altered the distribution pattern of LPL or GPIHBP1 in mouse pancreas. PMID: 23186339
- Findings indicate that miR-467b may regulate lipid accumulation and proinflammatory cytokine secretion in oxLDL-stimulated macrophages by targeting the LPL gene. PMID: 22963823
- ABCG1 controls LPL activity and promotes lipid accumulation in human macrophages in the presence of triglyceride-rich lipoproteins. PMID: 22772754
- variation in Lmf1 expression is a posttranslational determinant of LPL activity. PMID: 22345169
- lipoprotein retention in Bruch's membrane is mediated by lipoprotein lipase PMID: 21801873
- Hematopoietic cell-derived LPL could efficiently ameliorate severe hypertriglyceridemia and hypo-alpha-cholesterolemia at the compensation of increased triglyceride content of liver PMID: 21980507
- These results suggest that downregulation of miR-467b is involved in the development of hepatic steatosis by modulating the expression of its target, LPL. PMID: 21986524
- Uptake of dietary retinoids at the maternal-fetal barrier: in vivo evidence for the role of lipoprotein lipase and alternative pathways. PMID: 21795711
- LPL gene expression appears to be under dietary control: supplementation with cyanidin-3-O-beta-glucoside appears to up-regulate LPL in plasma and skeletal muscle but down-regulate LPL in visceral adipose tissue in the KK-Ay mouse model of diabetes PMID: 21360538
- the cleavage of ANGPTL4 by these PCs modulates its inhibitory effect on LPL activity. PMID: 21398697
- LPL is a novel Abeta-binding protein promoting cellular uptake and subsequent degradation of Abeta. PMID: 21177248
- Data show that apoC-II and LPL mRNAs correlate temporally and geographically with surfactant lipid synthesis in preparation for birth and suggest that fatty acid recruitment from the circulation by apoC-II-activated LPL is modulated by apoC-II secretion. PMID: 21059267
- PPAR-alpha response is generated by unbound fatty acids released locally by lipase activity and not by circulating plasma fatty acids. PMID: 20421589
- Results show that cotransfection of LPL with wild-type Lmf1 restores its ability to support normal lipase maturation. PMID: 19471043
顯示更多
收起更多
-
亞細胞定位:Cell membrane; Peripheral membrane protein; Extracellular side. Secreted. Secreted, extracellular space, extracellular matrix.
-
蛋白家族:AB hydrolase superfamily, Lipase family
-
組織特異性:Detected in white and brown adipose tissue and heart muscle, especially at the lumenal surface of capillaries. Detected on capillary endothelium in the lactating mammary gland. Detected in blood plasma (at protein level). Expressed in liver, epididymal fa
-
數據庫鏈接:
Most popular with customers
-
Human Transforming Growth factor β1,TGF-β1 ELISA kit
Detect Range: 23.5 pg/ml-1500 pg/ml
Sensitivity: 5.8 pg/ml
-
-
-
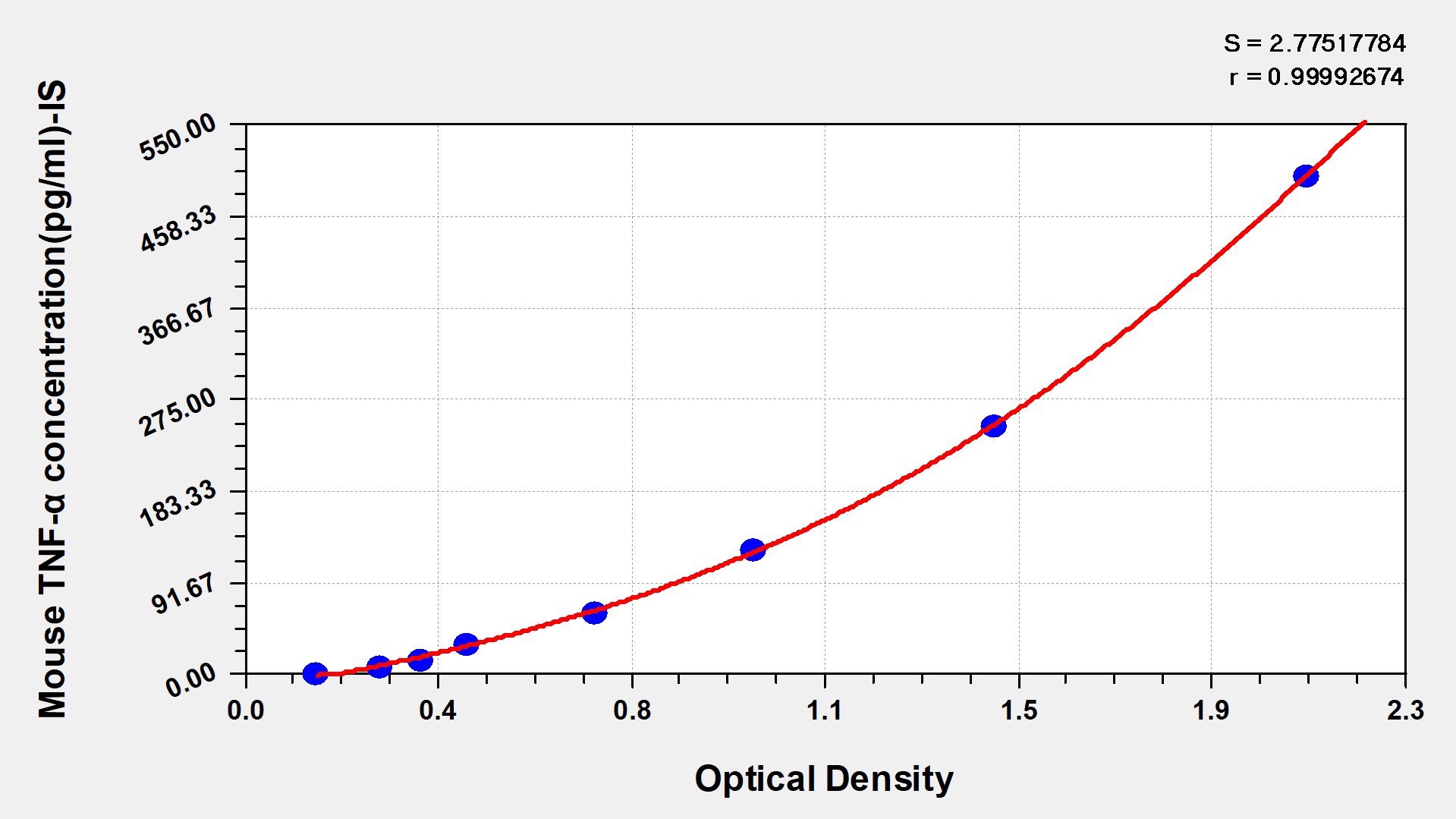
Mouse Tumor necrosis factor α,TNF-α ELISA Kit
Detect Range: 7.8 pg/ml-500 pg/ml
Sensitivity: 1.95 pg/ml
-
-
-
-