-
中文名稱:小鼠鐵調(diào)素(Hepc)酶聯(lián)免疫試劑盒
-
貨號:CSB-E14395m
-
規(guī)格:96T/48T
-
價格:¥3600/¥2500
-
其他:

產(chǎn)品詳情
-
產(chǎn)品描述:小鼠鐵調(diào)素(Hepc)酶聯(lián)免疫試劑盒(CSB-E14395m)為雙抗夾心法ELISA試劑盒,定量檢測血清、血漿、組織勻漿樣本中的HAMP含量。HAMP(Hepcidin/HAMP)是一種由肝臟產(chǎn)生的激素,主要調(diào)節(jié)鐵的吸收和分布。它通過促進鐵通道蛋白的內(nèi)吞和降解,減少鐵的吸收和釋放,維持鐵穩(wěn)態(tài)。HAMP的表達受多種因素調(diào)控,如系統(tǒng)性鐵負荷、炎癥因子、生長代謝信號等。研究HAMP的機制有助于開發(fā)治療鐵代謝相關(guān)疾病的新方法。試劑盒檢測范圍為0.78 ng/mL-50 ng/mL,本產(chǎn)品可廣泛應(yīng)用于鐵代謝調(diào)控機制研究、鐵相關(guān)疾病動物模型構(gòu)建或藥物干預(yù)實驗中Hepc動態(tài)變化的精準檢測本品僅用于科研,不用于臨床診斷,產(chǎn)品具體參數(shù)及操作步驟詳見產(chǎn)品說明書。
-
別名:Hamp ELISA Kit; Hamp1 ELISA Kit; Hepc ELISA Kit; Hepc1 ELISA Kit; Hepcidin ELISA Kit
-
縮寫:HAMP
-
Uniprot No.:
-
種屬:Mus musculus (Mouse)
-
樣本類型:serum, plasma, tissue homogenates
-
檢測范圍:0.78 ng/mL-50 ng/mL
-
靈敏度:0.195 ng/mL
-
反應(yīng)時間:1-5h
-
樣本體積:50-100ul
-
檢測波長:450 nm
-
研究領(lǐng)域:Cardiovascular
-
測定原理:quantitative
-
測定方法:Sandwich
-
精密度:
Intra-assay Precision (Precision within an assay): CV%<8% Three samples of known concentration were tested twenty times on one plate to assess. Inter-assay Precision (Precision between assays): CV%<10% Three samples of known concentration were tested in twenty assays to assess. -
線性度:
To assess the linearity of the assay, samples were spiked with high concentrations of mouse Hepc in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay. Sample Serum(n=4) 1:1 Average % 87 Range % 83-91 1:2 Average % 90 Range % 85-96 1:4 Average % 94 Range % 89-99 1:8 Average % 97 Range % 93-102 -
回收率:
The recovery of mouse Hepc spiked to levels throughout the range of the assay in various matrices was evaluated. Samples were diluted prior to assay as directed in the Sample Preparation section. Sample Type Average % Recovery Range Serum (n=5) 93 90-96 EDTA plasma (n=4) 99 94-103 -
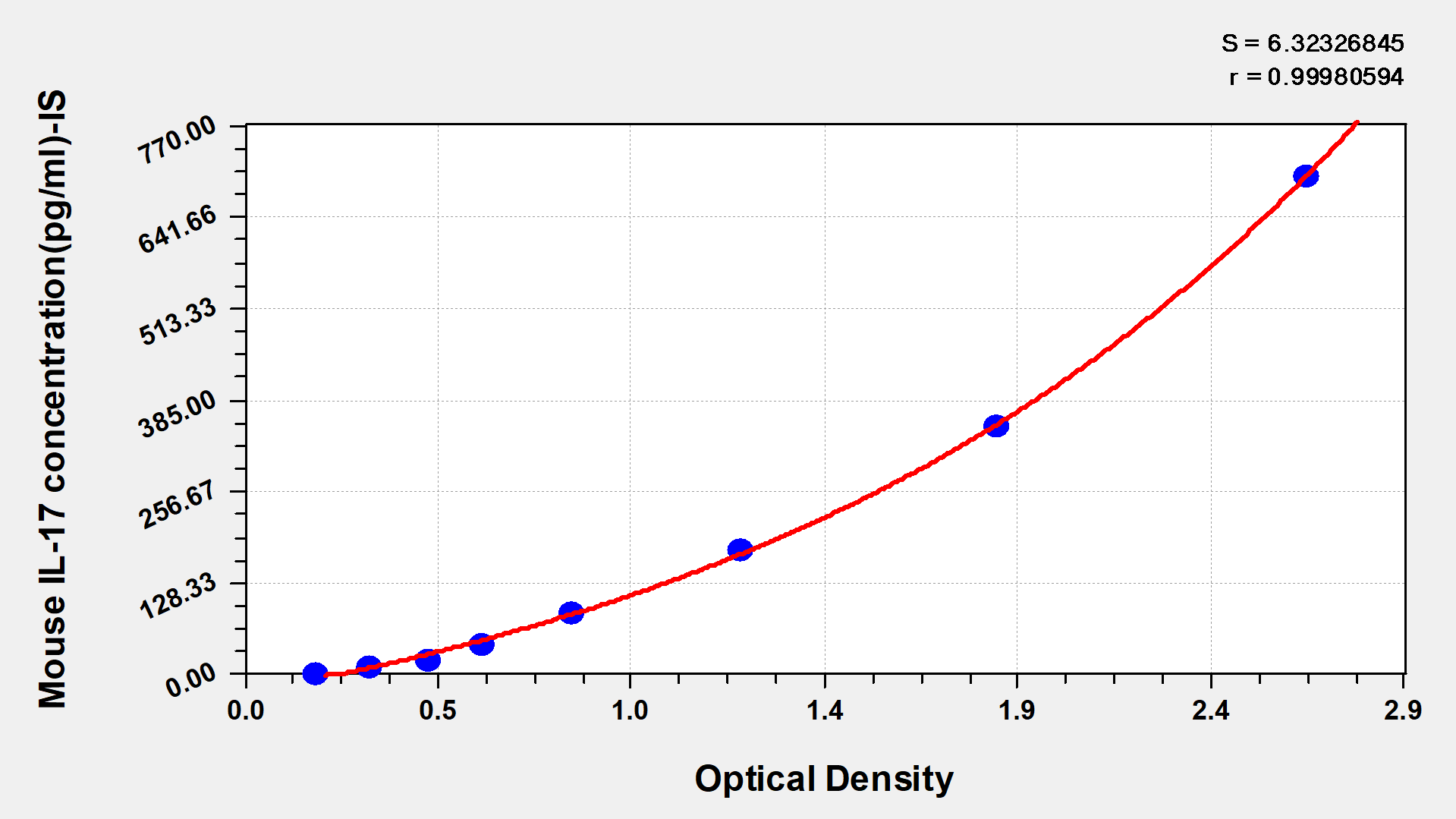
標準曲線:
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed. 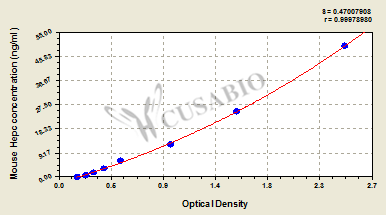
ng/ml OD1 OD2 Average Corrected 50 2.443 2.568 2.506 2.328 25 1.525 1.598 1.562 1.384 12.5 0.999 0.975 0.987 0.809 6.25 0.560 0.552 0.556 0.378 3.12 0.413 0.403 0.408 0.230 1.56 0.308 0.319 0.314 0.136 0.78 0.249 0.253 0.251 0.073 0 0.177 0.179 0.178 -
數(shù)據(jù)處理:
-
貨期:3-5 working days
引用文獻
- HIF-PHD inhibitor desidustat ameliorates iron deficiency anemia VJ Patel, A Joharapurkar, SG Kshirsagar, MS Patel, HH Savsani, HS Dodiya, MH Rakhasiya,Toxicology and Applied Pharmacology,2024
- Social defeat stress impairs systemic iron metabolism by activating the hepcidin–ferroportin axis E Kasahara,FASEB BioAdvances,2024
- Differences in DNA methylation of HAMP in blood cells predicts the development of type 2 diabetes M Ouni,Molecular metabolism,2023
- VHL-recruiting PROTAC attenuates renal fibrosis and preserves renal function via simultaneous degradation of Smad3 and stabilization of HIF-2α J Yang,Cell & bioscience,2022
- Hepcidin inhibition improves iron homeostasis in ferrous sulfate and LPS treatment model in mice V Patel,Drug research,2021
- Pharmacological inhibition of prolyl hydroxylase protects against inflammation-induced anemia via efficient erythropoiesis and hepcidin downregulation Jain M, et al,European journal of pharmacology,2018
- Hepatocyte toll-like receptor 4 mediates lipopolysaccharide-induced hepcidin expression .Yong-Soo Lee,et al,Experimental & Molecular Medicine,2017
- Pharmacological Characterization of ZYAN1, a Novel Prolyl Hydroxylase Inhibitor for the Treatment of Anemia. Jain MR.et al,Drug Res (Stuttg).,2016
產(chǎn)品評價

相關(guān)產(chǎn)品
靶點詳情
-
功能:Liver-produced hormone that constitutes the main circulating regulator of iron absorption and distribution across tissues. Acts by promoting endocytosis and degradation of ferroportin, leading to the retention of iron in iron-exporting cells and decreased flow of iron into plasma. Controls the major flows of iron into plasma: absorption of dietary iron in the intestine, recycling of iron by macrophages, which phagocytose old erythrocytes and other cells, and mobilization of stored iron from hepatocytes.
-
基因功能參考文獻:
- results indicate that a 0.25% carbonyl iron diet is sufficient to induce maximal hepatic hepcidin response. Importantly these results also demonstrate that in a chronic setting of iron administration, the amount of excess hepatic iron may not further influence hepcidin regulation and that expression of hepcidin plateaus at lower hepatic iron levels. These studies provide further insights into the regulation of this import PMID: 29752985
- Increased serum hepcidin contributes to the anemia of chronic kidney disease in a murine model. PMID: 27884972
- bone marrow transplantation between wild-type and TLR4 knockout mice revealed that hepatic TLR4-dependent hepcidin expression was comparable to macrophage TLR4-dependent hepcidin expression induced by LPS PMID: 29217822
- Hepc decreases in Cyp1b1-/- and gestational vitamin A deficiency mice resulted in stellate activation and lipogenesis suppression. PMID: 28583802
- data indicate that unlike with many other infections, hepcidin is decreased following M.tb infection, and show that hepcidin ablation does not influence M.tb growth in vivo PMID: 29324800
- Hepcidin expression involves epigenetic regulation by histone deacetylase 3. PMID: 28864822
- Serum hepcidin levels were measured by competitive ELISA in wild-type and Inhbb-/- mice at baseline and 4 hours after LPS challenge. Although Smad1/5/8 signaling is not activated by inflammatory stimuli in the absence of activin B, this has no impact on the induction of hepcidin expression. PMID: 27903526
- hepcidin mRNA upregulation depends on M1 macrophage polarization PMID: 27667162
- Our data provide evidence that the interplay of these two hormones could help improve the understanding of the pathogenesis of atherosclerosis and NAFLD. PMID: 29158088
- downregulation of hepcidin by siRNA increased iron uptake in bone and liver, which aggravated unloading-induced bone loss. PMID: 27686598
- The authors generated mice with cardiomyocyte-specific deletion of hepcidin, or knock-in of hepcidin-resistant ferroportin. They find that while both models maintain normal systemic iron homeostasis, the mice nonetheless develop fatal contractile and metabolic dysfunction as a consequence of cardiomyocyte iron deficiency. PMID: 27897970
- Acute tacrolimus treatment transiently increases hepcidin in wild-type mice. FKBP12 preferentially targets the BMP receptor ALK2. ALK2 mutants defective in binding FKBP12 increase hepcidin expression in a ligand-independent manner, through BMP-SMAD signaling. PMID: 28864813
- Hamp1 mRNA and plasma hepcidin levels are not good predictors of tissue iron levels, at least in males PMID: 28798083
- The lack of effect of erythropoietin on hepcidin expression in mask mice can not be explained by changes in erythroferrone synthesis, as splenic erythroferrone content increased after erythropoietin administration in both C57BL/6 and mask mice. PMID: 29073189
- these results characterise a new model of rapidly inducible hepcidin disruption, and demonstrate the critical contribution of hepcidin to the hypoferraemia of inflammation PMID: 27423740
- Hepatic gene expression of hepcidin is regulated in beta-thalassemia by ATOH8. PMID: 28405918
- Endogenous hepcidin and its agonist mediate resistance to selected infections by clearing non-transferrin-bound iron. PMID: 28465342
- the relationship between erythropoiesis and the candidate erythroid regulators, was examined. PMID: 28135344
- Smad1 and Smad5 have overlapping functions to govern hepcidin transcription. Moreover, erythropoietin and erythroferrone target Smad1/5 signaling and require Smad1/5 to suppress hepcidin expression. PMID: 28438754
- Genetic inactivation of hepatic angiocrine Bmp2 signaling in Stab2-Cre mice caused massive iron overload in the liver and increased serum iron levels and iron deposition in several organs similar to classic hereditary hemochromatosis; these changes were mediated by decreased hepatic expression of hepcidin, a key regulator of iron homeostasis. PMID: 27903529
- These studies indicate that drug-like minihepcidins have a potential as future therapeutics for untransfused beta-thalassemia and polycythemia vera. PMID: 27154187
- hepcidin induction by endoplasmic reticulum stress involves the central SMAD1/5/8 pathway PMID: 27483343
- Increased hepcidin in transferrin-treated thalassemic mice correlates with increased liver BMP2 expression and decreased hepatocyte ERK activation. PMID: 26635037
- The combined data presented here highlighted a crucial role of ERFE in regulating hepcidin expression and systematic iron homeostasis under phenylhydrazine-induced hemolytic anemia. PMID: 27067488
- The results suggest that physiologic hepcidin levels are insufficient to alter Fpn levels within the retinal pigment epithelium and Muller cells, but may limit iron transport into the retina from vascular endothelial cells. PMID: 26506980
- In TNF(DeltaARE/+) and IL-10(-/-)-mice hepatic hepcidin expression and protein content was significantly lower than in corresponding wild-types. PMID: 26302924
- In Angiotensin II treated mice, duodenal divalent metal transporter-1 and ferroportin expression levels were increased and hepatic hepcidin mRNA expression and serum hepcidin concentration were reduced. PMID: 25096756
- results clarify how commensal bacteria affect hepcidin expression and reveal a novel connection between IL-1beta and activation of BMP signaling. PMID: 26515063
- Activation of TLR4 signaling and NF-small ka, CyrillicB are involved in the suppression of hepcidin gene transcription by alcohol in the presence of inflammation in the liver. PMID: 25232250
- Study describes extensive blood vessel damage in Alzheimer's disease brain and a reduction in hepcidin and ferroportin levels PMID: 24252754
- Hepatic hepcidin plays an important role in sepsis through regulation of iron metabolism PMID: 25264597
- Hepc1(-/-) mice had a phenotype of low bone mass and alteration of the bone microarchitecture, most likely caused by a decreased osteoblastic activity PMID: 24652331
- Hepcidin mRNA expression was increased in the D-galactose (D-gal) group, decreased in the caloric restriction (CR) group, and was basically unchanged in the D-gal-CR group. PMID: 24044515
- results provide evidence that HFE induces hepcidin expression via the BMP pathway: HFE interacts with ALK3 to stabilize ALK3 protein and increase ALK3 expression at the cell surface. PMID: 24904118
- Data suggest that hepcidin-1 expression is up-regulated in obesity in visceral/subcutaneous adipose tissue (but down-regulated in liver); hepcidin may play local roles in modulating both iron metabolism and inflammation in adipose tissue. PMID: 24418880
- Hepcidin expression is upregulated in interleukin (IL)-10-deficient mice and downregulated in wild-type mice with dextran sulfate sodium-induced colitis. PMID: 24973448
- this study shows that the liver-specific KO mice fully recapitulate the severe iron overload phenotype observed in the total KO mice, with increased plasma iron and massive parenchymal iron accumulation. PMID: 24646470
- Results indicate a negative role of hepcidin in modulating liver regeneration. PMID: 24123375
- The Hamp1 and interleukin-6 knock out genotype resulted in improved erythropoiesis in aged mice. PMID: 23996485
- Hepcidin deficiency resulted in a marked reduction of bone load-bearing capacity likely through enhancing bone resorption, suggesting a direct correlation between hepcidin deficiency and bone loss. PMID: 24561287
- Hepcidin regulates intrarenal iron handling at the distal nephron. PMID: 23615502
- Data suggest that Hamp1 expression in liver can be regulated by dietary factors (here, down-regulated by a hematinic dietary supplement, black soyabean seed coat extract). PMID: 24387766
- Identify a link between glucose and iron homeostasis, showing that hepcidin is a gluconeogenic sensor in mice during starvation. PMID: 24361124
- Direct transcriptional suppression of hepcidin gene (HAMP) expression mediated by 1,25-dihydroxyvitamin D binding to the vitamin D receptor causes decrease in hepcidin mRNA levels PMID: 24204002
- The Brucella abortus model shows multifactorial pathogenesis of inflammatory anemia including iron restriction from increased hepcidin, transient suppression of erythropoiesis, and shortened erythrocyte lifespan. PMID: 24357728
- Knockout mice exhibited different patterns in the development and resolution of anemia, supporting the notion that interleukin 6 and hepcidin play distinct roles in modulating erythropoiesis in anemia of inflammation. PMID: 24357729
- Exogenous mouse IgG1 Fc fused to the N terminus of mouse IL-22 induced hepcidin production. Ab-mediated blockade of hepcidin partially reversed the effects on iron biology caused by IL-22R stimulation. PMID: 23836059
- Iron overload increased pro-hepcidin that remained high in hypoxia. PMID: 23656253
- A model is proposed that suggests that unlike proteases, which are irreversibly bound to activated alpha2M, hepcidin remains labile and available to down-regulate Fpn1. PMID: 23846698
- role of hepcidin in the setting of hypoferremia during acute inflammation PMID: 23637785
顯示更多
收起更多
-
亞細胞定位:Secreted.
-
蛋白家族:Hepcidin family
-
組織特異性:Highly expressed in the liver and to a much lesser extent in the heart. Secreted in blood.
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
Human Transforming Growth factor β1,TGF-β1 ELISA kit
Detect Range: 23.5 pg/ml-1500 pg/ml
Sensitivity: 5.8 pg/ml
-
-
-
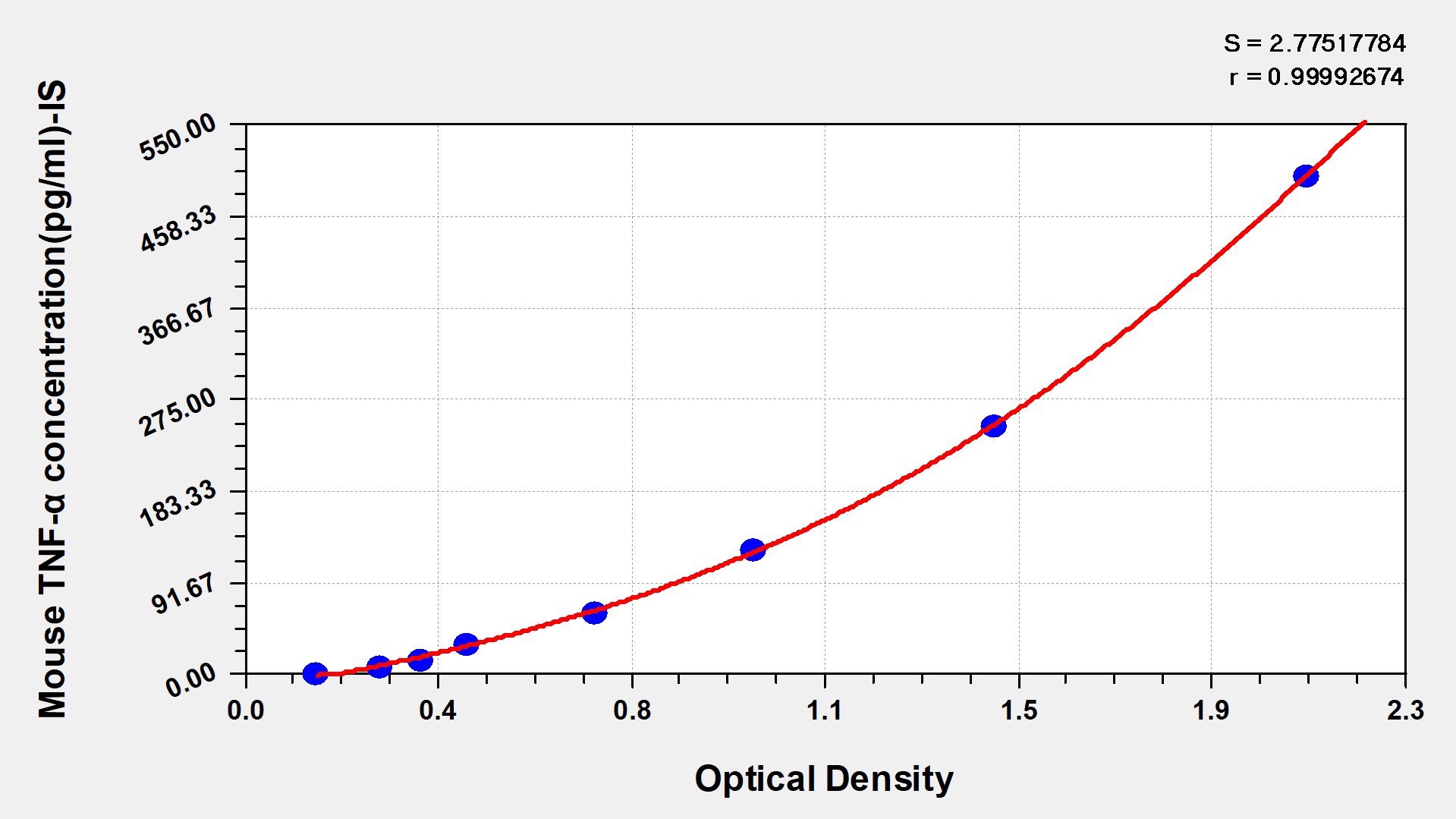
Mouse Tumor necrosis factor α,TNF-α ELISA Kit
Detect Range: 7.8 pg/ml-500 pg/ml
Sensitivity: 1.95 pg/ml
-
-
-
-